
Modern Black Sea oceanography applied to the end-Permian extinction event
2015-08-22 03:04:20SteveKershaw
Journal of Palaeogeography 2015年1期
Steve Kershaw
Institute for the Environment, Health and Societies, Brunel University,Kingston Lane, Uxbridge, Middlesex, UB8 3PH, UK
Abstract The modern Black Sea has a mixed upper layer in the top 150-200 m of the water column, below which the water is anoxic, separated from the mixed layer by a redox boundary. There is limited vertical movement of water. Pyrite framboids form in the water column of the anoxic zone, then have been traditionally interpreted to sink immediately and accumulate in the sediments of the Black Sea. Thus the occurrence of framboids in sediments in the rock record is widely interpreted to indicate poorly oxygenated to anoxic conditions in ancient environments. However, in the Permian-Triassic boundary (PTB) microbialites of South China, which formed in shallow marine conditions in contact with the atmosphere, the published occurrence of framboids is inconsistent with abundant gastropod and ostracod shells in the microbialite. Furthermore, in the modern Black Sea, (a) framboids may be suspended, attached to organic matter in the water column, thus not settle to the sea floor immediately after formation; and (b) the redox zone is an unstable complex area subject to rapid vertical water movement including occasional upwelling. The model presented here supposes that upwelling through the redox zone can lead to upward transport of suspended pyrite framboids into the mixed layer. Advective circulation could then draw suspended framboids onto the shelf to be deposited in oxygenated sediments. In the Permian-Triassic transition, if framboids were upwelled from below the redox boundary and mixed with oxygenated waters, sediment deposited in these conditions could provide a mixed signal for potentially misleading interpretations of low oxygen conditions. However, stratigraphic sampling resolution of post-extinction microbialites is currently insufficient to demonstrate possible separation of framboid-bearing layers from those where framboids are absent.
Key words mass extinction, Permian-Triassic boundary, microbialite, ostracods, gastropods, anoxia, Black Sea*
1 Introduction and aims
Causes of the end-Permian mass extinction remain problematic, but a growing body of evidence points to low oxygen levels throughout much of the ocean system. A range of measures of low oxygen includes pyrite framboids (Bond and Wignall, 2010; Tianet al., 2014),sulphur isotopes (Paytanet al., 2011), redox-sensitive elements (e.g., Algeoet al., 2010) and biomarkers (e.g.,Luoet al., 2013). Much work focuses on open shelf settings, and is establishing an interpretation of reduced oxygen in the deeper shelf and the deeper open ocean.However, there remains an issue of interpretation of oxygenation in locations where physical bio-sedimentary and shelly fossil evidence point to normal levels of oxygenation. For example, at Meishan, the GSSP(Global stratotype section and point) for the Permian-Triassic boundary, the open shelf sediments there contain abundant trace fossils, including open tunnels indicative of excavating organisms that must have needed oxygen.Danovaroet al.(2010) showed that modern metazoans can exist in low oxygen settings; these are very small metazoans in the Mediterranean Sea. Gingraset al.(2011) demonstrated that small metazoans can live in oxygenated microenvironments, receiving oxygen from photosynthesizing microbial organisms in otherwise low oxygen settings. These observations were explored by Forelet al.(2013) for the Permian-Triassic boundary (PTB) microbialites. However, those interpretations can apply only to situations where the amount of oxygen produced locally can be matched to the sedimentary evidence of shelly remains in the sediments. If there are abundant shelly remains, as in the case of many PTB microbialites, then local oxygen production within the microbialite may not have been sufficient to reasonably explain presence of abundant shelly fossils. There is also the issue, in the fossil record, of demonstrating whether or not the shelly remains were alive at the same time as the photosynthesizing oxygen-producing microbia. In the geological record, the issue of time-averaging in sediments is an important barrier to proving co-existence of organisms.
This paper addresses the issue of whether the microbialite system, that developed immediately after the end-Permian extinction event, grew in oxygenated conditions or not, using information from oceanographic processes in the modern Black Sea. The issue arises because of the discovery of pyrite framboids in the microbialite in one site (Laolongdong, Chongqing,South China; see Liaoet al., 2010). In general, the PTB microbialites are rich in shelly remains of ostracods and gastropods that may be reasonably viewed as requiring fully oxygenated conditions in which to grow.Therefore there is a seeming contradiction between the presence of pyrite framboids indicating low oxygen, on one hand, and shelly faunas in shallow marine waters indicating normal oxygen, on the other hand. Kershawet al.(2012) suggested that this contradiction could be explained by oxygen-poor water upwelling and choking the continental shelves. However, the current paper investigates this problem in relation to modern oceanographic processes in the Black Sea and provides an alternative potential explanation.
2 Black Sea oceanography
2.1 General pattern
There is a huge literature database on the Black Sea,because of its peculiar setting in the global aquatic system, being the only large water body currently existing that is largely anoxic. Critical to the current paper, the nature of the shallow Black Sea circulation is of great interest.
The Black Sea is divided into two basins, east and west, by a north-south orientated seafloor topographic ridge in its centre. However, water circulation consists of two forms of wind-driven water circulaton that influences both the upper (mixed) layer of oxygenated water, and the upper portion of the underlying anoxic water. Firstly,the surface waters are driven by westward-moving winds from the Caucasus Mountains in the northeast,to produce a cyclonic (anticlockwise) rim current along the continental slope (Figure 1), that varies from weak to strong, and completes one loop around the Black Sea in only a few months (Zatsepinet al., 2007). Secondly,two major anticyclonic gyres exist as permanent features,one in the east and one in the west basin (Figure 1). Also,anticyclonic mesoscale eddies and minor eddies develop across the Black Sea and are unstable, showing large changes over periods of only 3-6 months (Zatsepinet al., 2007; Tulzhikin, 2008).
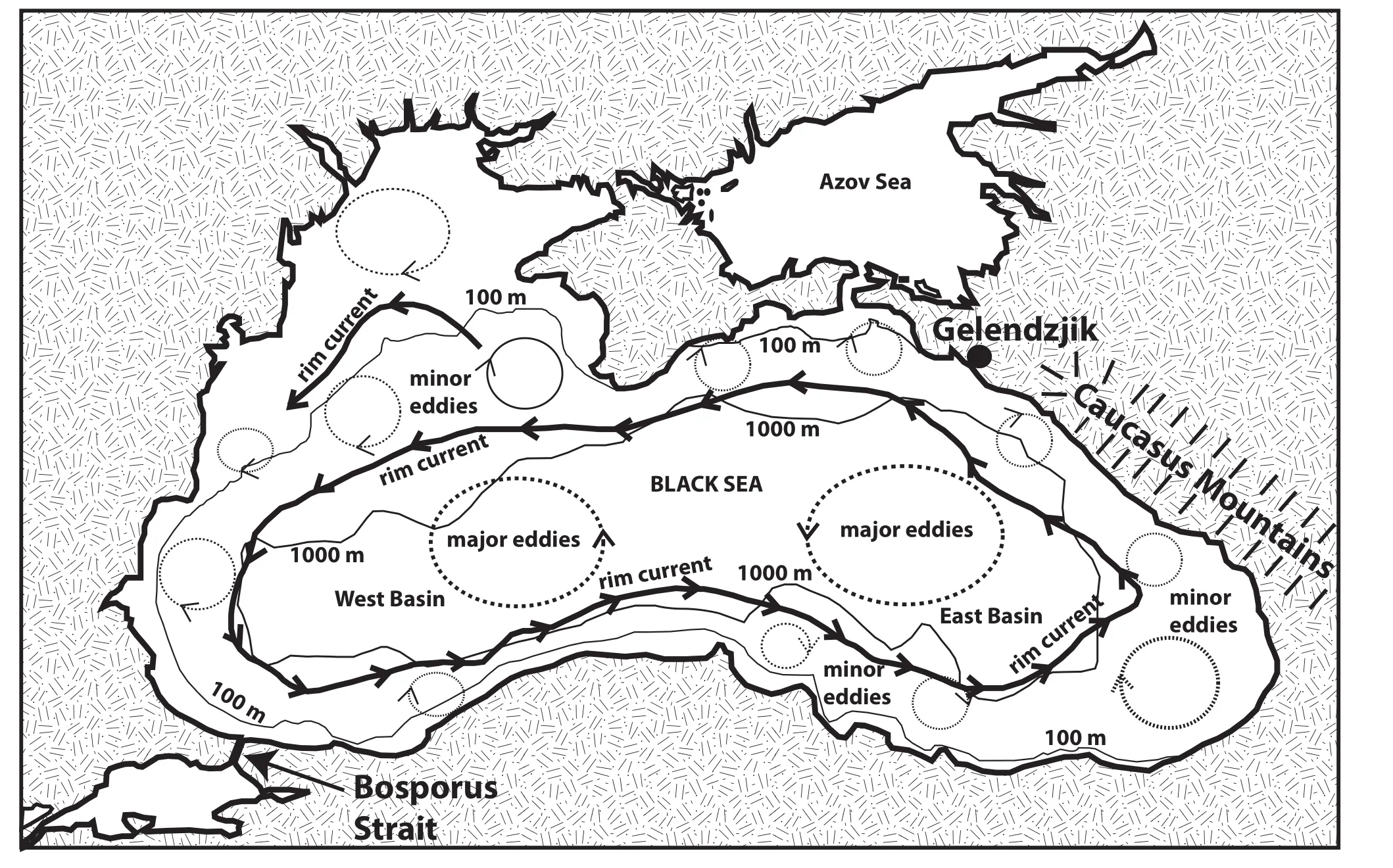
Figure 1 Map of the Black Sea showing the east and west basins, and the overall circulation patterns (based on Tuzhilkin, 2008; Fig.1). Note the Bosporus Strait in the southwestern margin, where dense warm saline water enters the Black Sea. This water sinks to an intermediate level and is transported across the Black Sea into the East Basin. Detailed oceanographic measurements by Ostrovskii and Zatsepin (2011) and sediment sampling by Ivanova et al. (2007) were undertaken offshore of Gelendzjik, on the northeastern coast,see text for details.
Thus, due to eddy flows, the upper 150-200 m of the Black Sea water mass is mixed by surface circulation and is aerated in contact with the atmosphere (Figure 2).Oxygen levels decline downwards, reaching zero at the redox boundary (Ostrovskii and Zatsepin, 2011) that lies between the upper mixed layer and the lower anaerobic waters which form most of the Black Sea mass. Volkovet al.(2007) demonstrated that the deeper water is composed of two distinct masses, and upwelling occurs from the lower mass. Major upwelling events occur occasionally in the Black Sea (Mihailovet al., 2012). Finally, Falinaet al.(2007) demonstrated that the dense saline water enter?ing the southwestern margin of the Black Sea via the Bos?porus (Figure 1) sinks to intermediate depths below the redox boundary and extends across into the east basin, as an intermediate water mass. Overall, there is considerable complex water movement in both the upper (mixed) and lower (anoxic) layers.
2.2 High-resolution short-term change
Ostrovskii and Zatsepin (2011) presented data col?lected daily over a week period, using a moored sub?mersible instrument platform (Aqualog) fixed to the sea floor off the shelf break in the northeastern Black Sea near Gelendzjik (Figure 1). Aqualog is automated to move up and down a fixed vertical rope over periods of a few hours, continuously recording data on whatever instruments are installed. Ostrovskii and Zatsepin(2011) recorded current strength, dissolved oxygen,fluorescence and turbidity (via acoustic backscatter),demonstrating vertical migrations of plankton within the mixed aerated layer, but critically showing that advective(horizontal) current flow in mesoscale eddies associated with easterly winds was dynamically variable and capable of transporting water onto and off the shelf areas in the mixed layer above the redox boundary. Upwelling and descending water masses occur uncommonly in the Black Sea, but is a recognized process of Black Sea oceanography (Ostrovskii and Zatsepin, 2011; Mihailovet al., 2012). Furthermore, the redox boundary shows vertical movements; as much as 40 m in one day has been observed (Alexander Ostrovskii, 2013; personal communication). Figure 2 shows a summary of short?term processes in the Black Sea mixed layer and upper parts of the anoxic layer.
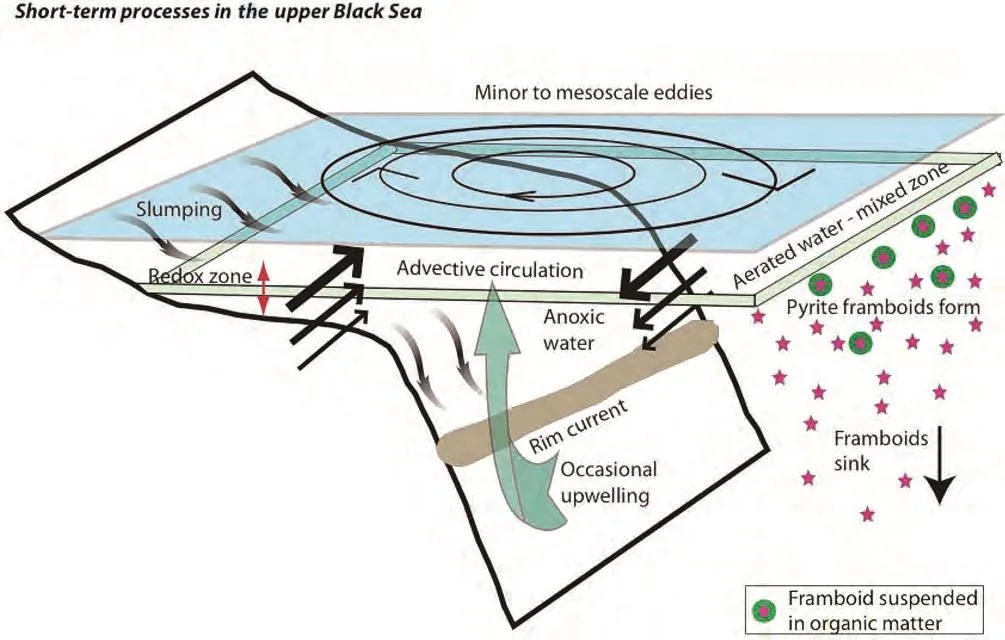
Figure 2 Diagram of conceptual short-term changes in the upper waters of the Black Sea. Wind-induced flows from the Caucasus Mountains east of the Black Sea drive eddies and the rim current that play an important part in dynamic motion of the Black Sea wa?ters. Slumping on the inclined substrate of the Black Sea is indicated. Eddy flow is advective (horizontal) and there is little vertical movement of water under normal conditions. However, occasional upwelling occurs in the Black Sea. Note also the redox zone and the formation of pyrite framboids. Published observations (see Wilkin et al., 1996; page 3898) of framboids suspended in the water on organic matter have important implications for the fate of pyrite framboids in sedimentary deposits; see text for discussion.
3 Observations from the Permian-Triassic boundary microbialites
Permian-Triassic boundary (PTB) microbialites con?tain abundant gastropod and ostracod faunas, between branches of microbial structure and in sedimentary layers within the microbialite (Kershawet al., 2012).Figure 3 shows part of the microbialite at the Dongwan site, Sichuan Province, South China, demonstrating the interrelationships between the microbial material and fossil?bearing sediments. Figure 4 and Figure 5 show de?tails of other samples from Dongwan, and draw attention to small scale changes within the microbialite, separating small depositional episodes, lithification and erosion in the shallow?water environments of microbialite growth. The implications of these detailed changes are significant to interpretation of environmental change within the micro?bialite unit, because there is clearly a temporal separation between successive parts of the microbialite. Current pub?lished information on pyrite framboids does not discrimi?nate these successive parts in detail.
4 A simple model for pyrite deposition in oxygenated environments
Data from long?term and short?term measurements in the modern Black Sea, cited above, demonstrate that the Black Sea processes are dynamic and complex at all scales. The occurrence of upwelling events provides the possibility of transport of suspended matter from the an?oxic layer upwards through the redox zone into the mixed layer. Although there is a generally agreed notion that py?rite framboids immediately sink to the sea floor after for?mation (e.g., Volkov and Neretin, 2008; Fig. 7), Wilkinet al.(1996; page 3898) reported that pyrite framboids occur suspended attached to organic matter in the Black Sea and in other places, such as Canadian lakes. These observations allow for the suspension of pyrite framboids in the water column that may be subject to vertical water movement.

Figure 3 Photographs of PTB microbialite at Dongwan, Sichuan, South China, showing shelly remains amongst microbial branches.a-Field photograph of microbialite domes and layered shell?rich sediment; b-Polished vertical section of microbialite and sediment;c-Dendritic branches of microbialite and fine-grained sediment. Note the shell remains between the branches. The scale unit is set to millimeter (mm).
If upwelled water carried suspended pyrite framboids up into the mixed layer, the possibility exists that advective currents could draw this water onto the shelf, and lead to the deposition of framboids in aerated sediments, see the model presented in Figure 6. When buried by active sedimentation on the shelf (from sediment precipitation,plus storm activity and slumping transporting sediment across the sea floor), framboid preservation is likely. Active sedimentation on shallow shelves during the Late Permian and Early Triassic is demonstrated by the sedimentary sequences illustrated in Figures 4 and 5, although slumping is not recorded in these ancient sediments, presumably due to low?angle shelves. Shelf sediment containing framboids in the rock record may thus lead to inaccurate interpretations of short?term events in the shallow?water sediments and influence models of ocean changes after the extinction.
5 Discussion
5.1 General features

Figure 4 Detail of stratigraphy of sediment and microbialite from the Dongwan site. The microbialite branches (M) are infi lled with sediment (S1), and then lithifi ed and eroded (E1). Three more layers of sediment are deposited, S2, S3 and S4, with lithifi cation and erosion (E2, E3) between them. This sample demonstrates the small scale complexity of one part of the microbialite, with important implications for investigations of oxygenation of the material. The scale unit is set to millimeter (mm).
The Black Sea is seen as a modern analogue for the ancient world ocean, particularly in the Paleozoic Era,where deep ocean waters have been considered as being low oxygen, under a different, less vigorous circulation re?gime, dominated by salinity controls (halothermal circula?tion), contrasting the modern processes where temperature is more important (thermohaline circulation; see Kershaw,2000 for discussion).
The model described above provides the possibility of depositing pyrite framboids, that formed in anoxic waters, into sediments formed on the shelf of the Black Sea above the redox zone. Ivanovaet al.(2007) reported angular unconformities in soft?sediment cores on the Black Sea shelf, also near Gelendzjik, where the Aqualog measurements were made. In soft sediments, such unconformities are best explained as a result of gravity?driven slumps, a common feature of the Black Sea margin.In the soft?sediment cores of maximum 10,000 years age, slumping would lead to some sediment mixing, but importantly provide the possibility of burying mobilized sediment after short?distance movement on the shelf.Pyrite framboids in these sediments should be preserved in sediment out of contact with the sediment/water interface,and therefore more likely to survive and be preserved permanently. Storm redistribution of sediment is also a potential mechanism to bury and preserve pyrite framboids.As yet the possible deposition of pyrite framboids in Black Sea mixed layer sediments has not been tested by sampling, but is an avenue for future investigation.
If this model is applied to the shallow seas after the end?Permian extinction event, it has the potential to explain how pyrite framboids could be present in the post?extinction microbialite along with shelly faunas of gastropods and ostracods. Indeed there are at least two possible explanations for the presence of framboids in the microbialite, both of which can be accommodated within the model described in this paper:

Figure 5 More detail of stratigraphy of sediment and microbialite from the Dongwan site. These photographs emphasize the erosion of the microbialite branches together with fi ne-grained carbonate sediment deposited between their branches, on a small scale, with subsequent sediment deposition and erosion. The scale unit is set to millimeter (mm).
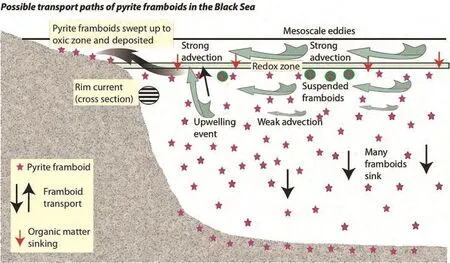
Figure 6 Cross?section through Black Sea showing an interpretation of the transport of pyrite framboids from the anoxic zone onto the shelf. Occasional upwelling draws suspended pyrite framboids into the upper mixed layer, where advective eddy circulation trans?ports the framboids to the shelf for deposition.
1) The framboids were uplifted from the anoxic zone in an upwelling event and any anoxic water also upwelled but mixed with aerated water and oxygenated. The framboids were then transported in oxygenated waters across the shelf to be deposited in shallow marine areas where the microbialites grew. For the Permian-Triassic boundary microbialite sequences, the illustrations in Figures 4 and 5 show layers of shelly fossils that may have been stormdeposited. In order for the framboids to be preserved, they would need to have been buried relatively quickly; the fine-grained sediment may be expected to have prevented oxygenated water causing oxidation of the framboids, and in cases where the microbialite layers immobilized the sediment (e.g., Wanget al., 2005) there is a strong case for preservation of framboids in low oxygen conditions beneath the redox boundary in the sediment.
2) The framboids were uplifted by upwelling as in “1)”,but the accompanying anoxic water retained its coherence,so that the framboids were carried in poorly oxygenated water and deposited in the microbialite. In this case, if the upwelling was a prolonged process that led to poorly oxygenated waters being a persistent feature of the shallow marine environments, then the possibility exists that the microbialites grew in poorly oxygenated conditions, a scenario suggested by Kershawet al.(2012).
Ostracod faunas associated with microbialites have been shown to reflect variations in oxygenated environments(Forelet al., 2009) with evidence of well-oxygenated and poorly-oxygenated waters in different places. Of interest,the pyrite framboids reported by Liaoet al.(2010) occur in the same locality (Laolongdong in Chongqing, South China) as lower oxygen conditions reported by Crasquin-Soleau and Kershaw (2005). The occurrence of abundant shelly remains of gastropods in PTB microbialites may then be accounted for by fluctuations of oxygenation of waters of the shallow shelf, so that the shelly faunas developed during periods of normal oxygenation between events of anoxic upwelling, as suggested by Kershawet al.(2012).Further evidence of oxygenation in the microbialites is in current work on redox-sensitive elements (Collinet al., 2012) and indicates that the microbialites grew in oxygenated conditions. Also, recent work by Tanget al.(2014) from the Chinese section at Yudongzi in the northwestern Sichuan Province, used abundant shelly faunas to indicate that the microbialites were oxygenated.Nevertheless, the possibility of fluctuations in oxygenation in that locality remains possible because a critical issue is the precision of sampling in the microbialite. Tanget al.(2014) sampled using a spaced resolution of 0.5 m per sample, leading to the possibility of omitting low-oxygen beds from the dataset, if they exist in the sequence. Clearly the way forward with such work is to sample at very high resolution, and not necessarily sample at fixed intervals,in order to ensure collection of all the changes in the sequence.
Liaoet al.(2010) sampled the microbialite at the Laolongdong site, Chongqing, and determined the presence of pyrite framboids to interpret lower dysoxic conditions,based on the diameters of the framboids. However, the precise sample locations of material for that study will have an important impact on the interpretations of framboids from Laolongdong. Unfortunately such details were not available, and therefore it is not possible in that study to discriminate between different beds within the microbialite. Nevertheless, the discovery of numerous samples that show consistency of pyrite framboid diameters (leading to the interpretation of dysoxia) is an important observation.
Liaoet al.(2010; Fig. 2) demonstrated that pyrite framboids of small diameters continued in the sediments above the microbialite, in the upper two samples. However,the fact that the microbialite disappeared from the section while the interpreted dysoxic conditions were maintained,suggests that dysoxia may not have been a principal control on the microbialite. Kershawet al.(2012) proposed that PTB microbialites were facies-controlled, and so the cessation of microbialite growth could be due to sea level rise, while the dysoxic conditions were maintained.Alternatively, using the model presented in this paper, if the framboids were all imported, then the entire logged section studied by Liaoet al.(2010) may have been in oxygenated sediments. Nevertheless, the results of Liaoet al.(2010) allow for the possibility that the microbialite material did not grow in dysoxic conditions, but that dysoxia was potentially due to temporary invasions into the shelf, between which the oxygenated environments allowed microbialites to grow. Note also that Knoll and Fischer (2011) drew attention to the logical deduction that shallow marine systems must be oxygenated because of the mixing of surface ocean waters in contact with the atmosphere. However, if anoxic upwelling events led to poorly oxygenated water transported across the shelf, the possibility exists that such waters could temporarily cut off the supply of oxygenated waters to the shallow sea floor,without the need for comprehensive invasion of anoxic water onto the shelf.
Details of the microbialite stratigraphy in examples in Figures 3-5 further emphasize the need for future highresolution work on PTB microbialites. Figures 4 and 5 are from adjacent samples at the same horizon and show how individual microbialite heads are filled with sediment, lithified and eroded, followed by further deposition,lithification and erosion. These two figures highlight the small-scale changes in sedimentary sequences that may have important implications for the environmental history of the rocks. Thus, the possibility exists that the microbialite and its enclosing sediment (M and S1 in Figures 4 and 5) formed in reduced oxygen conditions, but then were lithified and eroded prior to deposition of S2, S3 and S4 that contain rich shelly faunas and no microbialite. Thus,S2-S4 may have formed in well-oxygenated conditions and may even have been compiled by storm action,concentrating the shells.
Bond and Wignall (2010) indicated that euxinia in the end-Permian event could have occurred even above storm wave base. Nevertheless, it could be argued that such conditions could have been only temporary because of the fact that the upper ocean waters are in contact with the atmosphere, as noted by Knoll and Fischer (2011), so that the mixed upper waters could presumably not stay anoxic for long periods. Thus it is easy to visualise that the PTB microbialite sequences, as shallow marine features,grew in largely oxygenated conditions, and the presence of pyrite framboids was due to either transient lowoxygen conditions in the oceans, or import of framboids in oxygenated waters; but this idea needs to be tested in future work. Thus, overall, the model presented here points the way to focus future research to test the alternatives discussed above, but requires high resolution sampling to fully investigate.
5.2 Oxygenation, ocean circulation and palaeogeography
While evidence of low-oxygen conditions occurs in many places across the Permian-Triassic boundary, some sites show no evidence, such as the Wasit Block in Oman where abundant and diverse faunas occur above the extinction level (Baud and Bernecker, 2010). Furthermore,there is clear evidence of palaeogeographic variations in structure of post-extinction microbialite communities(Kershawet al., 2012). Normally the upper waters would be expected to be well-circulated by gyres in Tethys, as modeled by Kidder and Worsley (2004). Eastern Tethys sites in the South China Block contain microbialites that are extensively recrystallized but contain remnant structures dominated by renalcid-type calcimicrobes (Kershawet al., 2012). In contrast, in western Tethys sites, principally in Turkey and Iran, the microbialites are composed of sediments, as stromatolites (laminated and layered clotted micrites) and thrombolites (thromboidal clotted micrites),see Figure 7. Figure 7 also draws attention to western Pangaea, where PTB microbialites have not been found,despite that the area being in the right latitudes for microbialite growth, given the low latitude abundance of microbialites in Tethys. The reasons for such profound differences are currently uknown and require further investigation.
Bond and Wignall (2010) surveyed pyrite framboids in different locations across the Permian-Triassic boundary,as follows:
Western Tethys: In Hungary, although framboids were found in the boundary shale directly below the microbialite, none were found in the overlying stromatolites, in a ramp setting. In Italy, there is little microbialite preserved(Mazzin Member) with rare framboids only at the base of the microbialite, yet framboids were common in the underlying Tesero oolite, which is a mobile sediment subject to strong water movement. Although Bond and Wignall(2010) interpreted the Tesero oolite to have been deposited in a time when anoxic water expanded into shallow depths,the possibility of inwashed framboids to be deposited in oxygenated conditions, then buried, as proposed in the model presented in Figure 6, cannot be ignored.
Eastern Tethys:The South China sequences studied by Bond and Wignall (2010) are all open shelf settings,and contain some framboids. The only microbialite site so far recorded with framboids is Laolongdong (Liaoet al.,2010).
Even though there are geographic differences between microbialites of the various sites in Tethys, framboid occurrences in both eastern and western Tethys demonstrate that upwelling was a common feature of the ocean circulation, yet there were barriers to horizontal migration of microbial organisms across Tethys, suggesting that mixed layer advective motion may not have been as effective as in modern oceans. In the modern Black Sea, advection is very effective in moving water around the Black Sea surface waters, as are the open ocean gyres. Thus the contrasts between microbialites will perhaps force reassessment of ocean circulation interpretations of the PTB times,and emphasize the key role that they play in understanding processes across this important episode of Earth history.
6 Conclusions
Modern Black Sea oceanography is applied here to propose a model that pyrite framboids may have been upwelled into the upper mixed layer of the Black Sea and transported onto the shelf by advective eddies to be deposited in oxygenated sediments. This model needs to be tested by sampling oxygenated sediments of the Black Sea shelf and search for pyrite framboids.

Figure 7 Simplified palaeogeography of the Permian-Triassic, with major features of microbialite geographic distribution. Note the major differences of microbialite composition between western and eastern Tethys, and the lack of microbialites in western Pangaea, in latitudes and locations where they would be expected to occur. See Kershaw et al. (2012) for more details of the geographic distribution of the microbialites.
In the Permian-Triassic transition, the possibility of deposition of pyrite framboids in oxygenated sediments may solve the contradiction of co?occurrence of framboids and shelly faunas in the microbialites; this model therefore has profound implications for interpretations of ocean oxygenation of the Permian-Triassic transition.
Palaeogeographic differences between microbialites of the various areas within Tethys imply that upper ocean circulation may not have been effective in the Permian-Triassic transition, in contrast to modern oceans, including the Black Sea, where advective circulation is very effective for migration of biota.
This work, and in particular the uncertainties demon?strated in the discussion, shows the great need for future careful fieldwork and high-resolution sampling of Permi?an-Triassic boundary sequences in order to develop the ideas presented in this paper.
Acknowledgements
I am most grateful to scientists at the P. P. Shirshov Insti?tute of Oceanology, Moscow, for support and discussions during research visits to the Black Sea in 2012 and 2013. I particularly thank Alexander Ostrovskii for discussion on Black Sea processes and am indebted to Victoria Putans,Nikolai Esin and Kolya Esin for logistic support. Max McGuire, Noel Healy, Maria Zhdan and Tanya Zhdan are thanked for their support in Russia. Russian work was sup?ported by EU?funded project PIRSES?GA-2009-247512.Prof. Zeng?Zhao Feng and Prof. Yong?Biao Wang are thanked for their comments on an earlier draft. This paper contributes to: (a) project “CLIMSEAS”: Climate Change and Inland Seas: Phenomena, Feedback and Uncertainties,the Physical Science Basis; and (b) project IGCP630:“Permian?Triassic climatic and environmental extremes and biotic response”.
 Journal of Palaeogeography2015年1期
Journal of Palaeogeography2015年1期
- Journal of Palaeogeography的其它文章
- Provenance and drainage system of the Early Cretaceous volcanic detritus in the Himalaya as constrained by detrital zircon geochronology
- 3D palaeogeographic reconstructions of the Phanerozoic versus sea-level and Sr-ratio variations
- Nonmarine time-stratigraphy in a rift setting: An example from the Mid-Permian lower Quanzijie low-order cycle, Bogda Mountains, NW China
- A revised Mississippian lithostratigraphy of County Galway (western Ireland) with an analysis of carbonate lithofacies, biostratigraphy, depositional environments and palaeogeographic reconstructions utilising new borehole data
- General regulations about submitting manuscripts to Journal of Palaeogeography
- 2nd International Palaeogeography Conference October 10?13, 2015