
Stem radialgrow th indicate the optionsof species,topography and stand management for artificial forests in thewestern Loess Plateau,China
2019-07-10 09:53:22ShengChunXiaoXiaoMeiPengQuanYanTianGongZhu
ShengChun Xiao,XiaoMeiPeng,QuanYan Tian,2,Gong Zhu
1.Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences(CAS),Lanzhou,Gansu 730000,China
2.University ofthe Chinese Academy of Sciences,Beijing 100049,China
3.Landscape Engineering DepartmentofSouth-North Mountain ofLanzhou City,Lanzhou,Gansu 730000,China
ABSTRACT An understanding of the differences in artificial forestbetween tree species,slope aspects,andmanagementoptions in arid environments is fundamentally important for efficientmanagementof these artificial systems;however,few studies have quantified the spatial and temporal differences in stem radial grow th of trees in the arid western Loess Plateau of China.Using dendrochronology,we assessed the grow th of threewoody species(the native shrub Reaumuria soongorica,the exotic shrub Tamarix ramosissima and tree Platycladus orientalis)by measuring the annual stem radial increment.We also describe the long-term grow th trends and responses to climatic factors on slopeswith differentaspects during periodswith and without irrigation.We found that precipitation during themain grow ing season was significantly positively correlated with ring grow th for all three species and both slope aspects.In addition,supplementalwater(e.g.,irrigation,rainwater harvesting)greatly relieved drought stress and promoted radial grow th.Our results suggest that as themain afforestation species in the Loess Plateau used for soil and water conservation,P.orientalis ismore suitable than T.ramosissima under rain-fed conditions.However,a landscape that combined a tree(P.orientalis)with a shrub(R.soongorica)and grassland appears likely to represent the bestmeansof ecological restoration in the arid western Loess Plateau.
Keywords:dendroecology;tree-ring;artificial forest;Loess Plateau
1 Introduction
China's Loess Plateau has experienced severe soil erosion,vegetation degradation,and desertification(Jiao et al.,2011).Since the late 1950s,a series of government-backed projects,including extensive tree planting,watershed erosion-controlmeasures,and the"Grain-for-Green"program,have been implemented to control soil erosion and restore vegetation(Chen etal.,2008;Zhang et al.,2009;Wang et al.,2011;Jiang et al.,2015).Under these programs,mostof the traditional farm land on steep slopes has been converted into grasslands,shrub-land,and forests during these restoration processes(Yang et al.,2012).As a result,erosion has been reduced or elim inated in about24%of the area where this problem formerly occurred,and vegetation cover has increased by 6%to 8%in forests of the Loess Plateau(Chen etal.,2008;Jiang etal.,2015).
However,the massive afforestation program has produced negative impacts on the environment,such as low shrub and tree survival rates and soil desiccation due to excessivewater consumption by the planted vegetation(Jiang et al.,2015).Although some trees and shrubsmay grow well soon after planting,their grow th ratemay decline significantly during later stages,and the plantsmay even die during drought years.Both local and external factors,including climate change,drought,introduction of unsuitable species,inappropriate planting densities,and a failure to account for the effects of the slope aspect,has led to soil desiccation and vegetation degradation in many areas;the ultimate resultof these processes has been a decline in the water generation and soil conservation functions of the artificial vegetation(Li et al.,2007;Chen et al.,2008;Fu et al.,2012;Jia and Shao,2013).The conflict between soil desiccation and the sustainability of vegetation has become increasingly important on the Loess Plateau(Liu and Shao,2015).Many studies have focused on the soil moisture regime in artificial forests(Li et al.,2007;Chen et al.,2008;Fu etal.,2012;Jia and Shao,2013)and on water consumption by individual trees(Yu et al.,2009),but few researchers have studied plant grow th,which is a key component of the soil-plant-atmosphere continuum(Xiao etal.,2015).
Long-term data is needed to better understand these interactions,and the annual grow th rings of woody vegetation can provide such informationwhenmore directmeasurements are unavailable(Xiao et al.,2015).Dendrochronology aims to establish long-term relationships between radial grow th and climatic variables(Fritts and Shatz,1975),and to assess how forestmanagementoptions influence the grow th of naturaland artificial forests(M?ren et al.,2015;To?go et al.,2015)on slopeswith differentaspects(Xiao et al.,2015)and in response to anthropogenic disturbances(Baldauf et al.,2015).Tree rings provide precise information on past tree grow th and its response to environmental changes(Spiecker,2002).Through analysis of the relationship between climate and tree grow th,dendroecology can provide insights into factors that lim it tree grow th(Antos et al.,2008).Many dendrochronological studies have focused on natural forests at high elevations in rocky mountainous areas on the Loess Plateau(Liu et al.,2013;Song et al.,2013;Fang et al.,2015;Li etal.,2015),but few researchers have studied artificial forests(Xiao etal.,2015).
Platycladus orientalis(Cupressaceae)is an evergreen native tree in China that is also naturalized in Korea,Japan and Iran(Jiang etal.,2015).Tamarix ramosissima(Tamarix)is a large shrub and is distributed commonly in North China and the arid regions in m iddle Asia(Liu,1987).P.orientalis and T.ramosissima were considered to be suitable woody species for afforestation on the Loess Plateau because of their good performance,including high post-transplanting survival,rapid early grow th,a well-developed root system,and a pleasant appearance in the landscape(Yu etal.,2009;Jiang etal.,2015).Reaumuria soongorica is a common native dwarf shrub in arid regions of China,including the plateau(Xiao et al.,2006;Xiao et al.,2015).On the western Loess Plateau,there are several different and relative older artificial forests of P.orientalis and T.ramosissima with varied slope aspect and managementmeasures.It provides relative abundant and comparable materials for our study,including the native dwarf shrub and two speciesof the artificial forests.
In this paper,we aimed to assess the grow th performance and adoptability of the three species on the differentm icroenvironment and management options on the arid western Loess Plateau.These results would provide guidance for ecological rehabilitation and artificial forestmanagementof this and sim ilar regions.We also hypothesized that(1)the stem grow th of the two afforestation species(P.orientalis and T.ramosissima)and the native dwarf shrub(R.soongorica)would be restricted by precipitation in this arid region,(2)this restriction would be relieved by supplemental water(irrigation or water harvesting),and(3)the three specieswould have different grow th performance and responses to the slope aspect andmanagement options.To test these hypotheses,we used dendrochronological methods to assess the annual stem radial increment of the three woody species,their long-term grow th trends,and their response to climatic factors on slopeswith differentaspects during periodswith and without irrigation.
2 Study area,m aterials and m ethods
2.1 Study area
The Loess Plateau is located in the upper andm iddle reaches of the Yellow River,between the western Taihang,eastern Riyue-Helanns,northern Qinling,and southern Yinsan mountains(100°54′E to 114°33′E,33°43′N to 41°16′N).The plateau covers a total area of 6.29×105km2,with an elevation of 1,200 to 1,600 m above sea level,and has a loess coverwith a thickness thatgenerally ranges from 30 to 80m(Zhu etal.,1983).This region is a transitional zone between the southeastern hum id monsoon climate and the northwestern continentaldry climate(Chen etal.,2008).
Our study was conducted north and south of Lanzhou City,in China's Gansu Province(Figure 1).The city is located in the western part of the Loess Plateau,which has a sem i-arid temperate climate.The Yellow River flows through the city.Annual mean rainfall averages 249.0±59.1 mm(1961 to 2013),with more than 84%falling from May to September.Annual pan evaporation(Φ20 cm)ranges from 1,400 to 1,800 mm.The mean annual air temperature is 7.2±0.6°C,with a mean monthly temperature of 20.6±1.2°C in July,and ameanmonthly temperature of-8.9±1.2°C in January(Xiao etal.,2015).The soil is a silt loam of loess origin,and can be classified as a Calciorthid(Li etal.,2006).The study area is located in a transitional zone between typical steppe and desert steppe vegetation,with low species richness and sparse vegetation composed of xerophytes and halophytes(Song etal.,2013).
2.2 Materials and methods
We selected three sampling sites for this study(Table 1).The G site is located south of Lanzhou,and includes two topographic situations(slopes with south and north aspects)and two species(P.orientalis and T.ramosissima).T.ramosissima seedlings(2-year old)were planted in the late 1950s and P.orientalis seedlings(3-to 4-year old)were planted in the early 1980s on flatmanmade terraces that were 1.5 to 6.0 m w ide and ran perpendicular to the slope.T.ramosissima were planted only at the edges of flat terraces,ata spacing of 2 m within the rows.P.orientalis were planted on the flat terracesata spacing of 2m within rowsand 2m between rows.The terraces were irrigated by broadcast spraying from a water pipeline before 1988,then were abandoned and only received water from precipitation.Mostof the T.ramosissima shrubs appear to be significantly damaged,with many dead branches,whereas P.orientalis outwardly appearnormal.
The X site is located in northern Lanzhou,in a local public park with flat topography.P.orientalis were planted along the road in the late 1950s and irrigated through furrows.A fter periodic pruning of every year,the trees remained just one trunk with 2 m height under branch.The L site was the native dwarf shrub R.soongorica,on a hill top in the northern part of the park,and received only natural precipitation.This site showed spontaneous recovery of vegetation in abandoned farm land after 30 years.
Due to the presence ofmultiple branches,eccentricity,and the short main trunk of the shrubs,sample discs were obtained at the soil surface at the L and G sites.At those sampling site,we sampled the largest stem from each shrub,including some dead but big stems,and obtained cross-sections at their root collars.We obtained cores of P.orientalis at 40 cm above the ground at the X site.Ateach site,more than 20 shrubs were selected for disk sampling,ormore than 20 cores were obtained from the large P.orientalis trees at the X site.Allsampleswere collected in the fallof 2014.
Follow ing standard dendrochronologicalmethods,we air-dried the sample discs and the groove-mounted core samples for several days,then planed and sanded their surfacewith progressively finer sandpaper to enhance the visibility of the boundaries between grow th rings.Tree rings in each disk were visually cross-dated along two radii.False andm issing ringswere easily identified by comparing these two radii(Xiao et al.,2007,2012,2015).Ring w idths were then measured to the nearest0.001mm using the LINTAB treering measurement system(http://www.rinntech.de).Because of asymmetries in the stems,we measured rings along both the largest and the smallest radii of the shrub or tree.Themeasured tree-ring sequences at each sampling site were cross-dated by a variety of means,including comparison based onmorphological characteristics that were clearly visible in the tree rings under am icroscope,matching the ring-w idth sequences and marker years among samples,and quality-checking the results with the COFECHA software(Holmes,1983;Grissino-Mayer,2001;Zhang et al.,2012).A few of the sequenceswere too short,and we did notuse them for further analysis.
A ring-w idth chronology for each species at each site was developed using the ARSTAN software(Cook and Holmes,1996).Using the mean grow th curve for a species at each site,we removed the effects of age-related grow th trends using a horizontal line based on the mean ring w idth or a straight line with negative slope.The detrended ring-w idth sequences were then averaged by year for the different samples from a given site.The resulting tree-ring chronologies represent grow th variations caused by common environmental forcing at the site.We calculated chronological statistics,such as themean sensitivity(a measure of the annual variability in treerings),mean inter-serial correlation(ameasure of the magnitude of the common signal among tree-ring sequences),the expressed population signal(EPS,which is a measure of the strength of environmental signals expressed by tree rings),subsample signal strength(SSS,which is a measure of the strength of environmental signals contained in a subsample of tree rings relative to the whole number of samples),the all-seriesmean correlation among all tree-ring serieswithin a population(Rbar,which is independent of sample size)(Briffa,1999),and the variance in the first principal component(PC1)of the tree-ring series(which is ameasure of the proportion of the signal in the tree rings that could be represented by the first principal component)to describe basic characteristics of the tree ring chronologies(Cook and Kairiukstis,1990;Zhang etal.,2012).
We compared mean raw ring w idths(Difference tests)between different species,different slope aspects,and periodswith and without irrigation(before and after 1988)to clarify grow th conditions under differentmanagementoptions ateach site.We examined the relationship(Pearson's r)between ringw idth chronologies(ring w idth index)at different sampling sites.We also exam ined the relationships between climatic factors(precipitation and temperature)and radial stem grow th by calculating the correlations between these factors(Pearson's r)and ringw idth chronologies,and between the irrigation and without-irrigation periods at the G sampling site,and also the periods of before and after 1988 at the X sampling site.For climatic variables,we chose the mean air temperature and total precipitation in each month from April to October of the current year,and total precipitation from M ay to August during the grow ing season.Climate data was obtained from the Gaolan meteorological station(36°21′N,103°56′E,1,690 m a.s.l.),north of Lanzhou City and about 10 km from the study sites.
3 Resu lts
3.1 Stem radial grow th characteristics
The tree's radial grow th curves varied among the species and sampling sites(Figure 2).The tree's grow th of P.orientalis at the GNP and GSP sites followed a parabolic curve(Figure 2a;site code refer to Table 1 and as follows),with peak grow th in them idto late 1990s,and localm inima for grow th rate during the late 1980s and 2000s.At the X0Psite,this species showed a long term decrease in grow th(Figure 2c).At the GNT and GST sites,T.ramosissima grow th showed an overall decreasing trend(Figure 2b),but showedmore than fourmajor fluctuations between increasing and decreasing grow th.T.ramosissima shrubs and P.orientalis trees showed sim ilar,simultaneous grow th decreases on slopes with north and south aspects when the irrigation stopped in 1988.The stem radial grow th trend of R.soongorica shrubs remained relatively constant throughout the study period(Figure 2c).
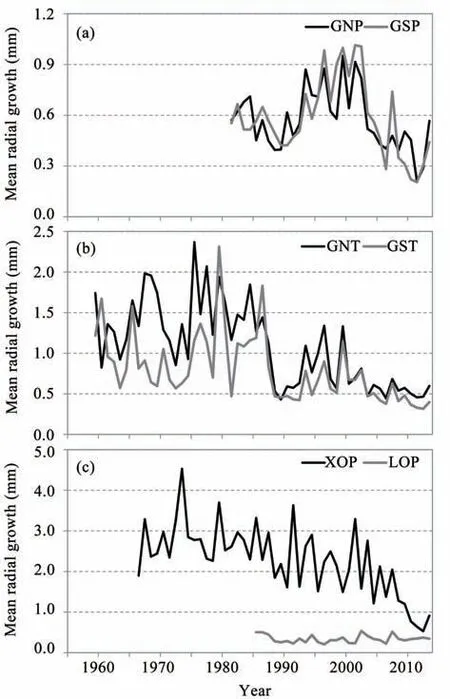
Figure 2 Mean stem radialgrow th curves for the three woody speciesat the sampling sites.A:P.orientalis on GNPand GSPsite;B:T.ramosissima on GNT and GST site;C:P.orientalis on X 0Psite and R.soongorica on L0Psite
Figure 3 compares the mean ring-w idth of the three species at the different sampling sites,with different slope aspects,and the T.ramosissima grow th curve during periodswith and without irrigation at the GNT and GST sites.Themean ring w idth of P.orientalis trees at the X0P site(2.28±0.84 mm,mean±SD)was significantly greater(P<0.05)than its grow th at the GNP(0.57±0.18)and GSP(0.58±0.22)sites,than T.ramosissima grow th at the GNT(1.08±0.50)and GST(0.80±0.41)sites,and greater than R.soongorica grow th at the L0R site(0.34±0.09)(Figure 3a).The mean ring w idth of P.orientalis at the X0Psite,where furrow irrigation was provided throughout the study period,was about four times thatat the GNPand GSP sites.Themean ring w idth of T.ramosissima was significantly greater(P<0.05)than that of P.orientalis at the same site.Themean ring w idth of T.ramosissima on the slopewith a north aspect(GNT)was significantly greater(P<0.05)than that on the slopewith a south aspect(GST).For the periodswith and without irrigation(Figure 3b),themean ring w idth of T.ramosissima during the irrigation period(GNTi and GSTi)was significantly greater(P<0.05)than that during the period without irrigation(GNTn and GSTn).During the irrigation period,themean ring w idth of T.ramosissima on the slope with a north aspect(GNTi)was significantly greater(P<0.05)than that on the slopewith a south aspect(GST).However,T.ramosissima grow th without irrigation did not differ significantly between the two slope aspects(GNTn and GSTn).
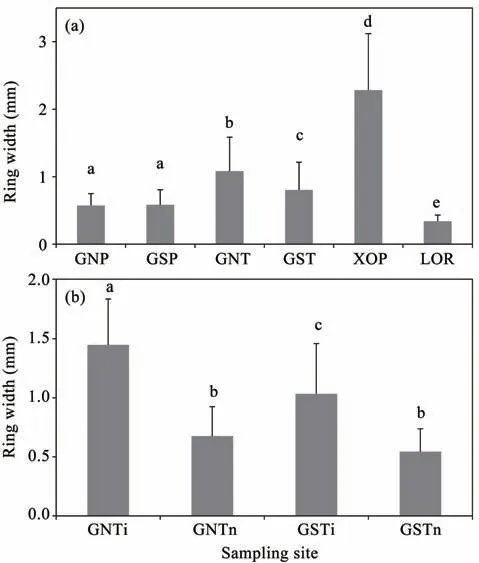
Figure 3 Average ringw idth of(a)the three speciesat the six sampling sites in GNP,GSP,GNT,GST,X 0Pand L0P,and(b)of T.ramosissima during the periodswith(GNTiand GSTi)and without irrigation(GNTn and GSTn)at the GNT and GST sites.Valuesaremean±SD.Bars fora given parameter labeledwith different lettersdiffersignificantly between the species,sampling sites,slope aspectsandmanagementoptions(P<0.05)
3.2 Characteristics of ring-width chronologies
A fter removal of the age-related radial grow th trends,we established six standard ringw idth chronologies(Figure 4).Table 2 summarizes the statistics for the standard chronologies at the six sampling sites.The chronologies extended from 1982 to 2013 at the GNP and GSP sites,from 1959 to 2013 at the GNT and GST sites,from 1966 to 2013 at the X0P site,and from 1985 to 2013 at the L0R site.The value of SSS reached 0.85 in 9,5,7,5,7,and 9 ring-w idth series in 1983,1982,1967,1979,1968,and 1987 at the GNP,GSP,GNT,GST,X0P,and L0R sites,respectively.The mean serial correlations were relatively high,except for values of 0.145 and 0.182 at the L0R and GNPsites,respectively.Themean sensitivity of the standard chronologies ranged from 0.249 to 0.335,and the proportion of the explained variation in the f i rst principal component ranged from 38.6%to 54.1%.During the common period for the standard chronologies,the EPS valueswere all greater than 0.90,and Rbarranged from 0.322 to 0.503.Due to the short time span and the credible values of those parameters,we used whole chronologies in our subsequentanalyses.
We also calculated the correlations among the six chronologies,and found that two species(P.orientalis and T.ramosissima)at the same sam pling site(G)had significantly correlated chronologies(P<0.05,Table 3).The chronology at the X0P site was only significantly correlated with that at the GSP site(r=0.384,P<0.05).The chronology at the L0R sitewas not significantly correlated with that of any other site.
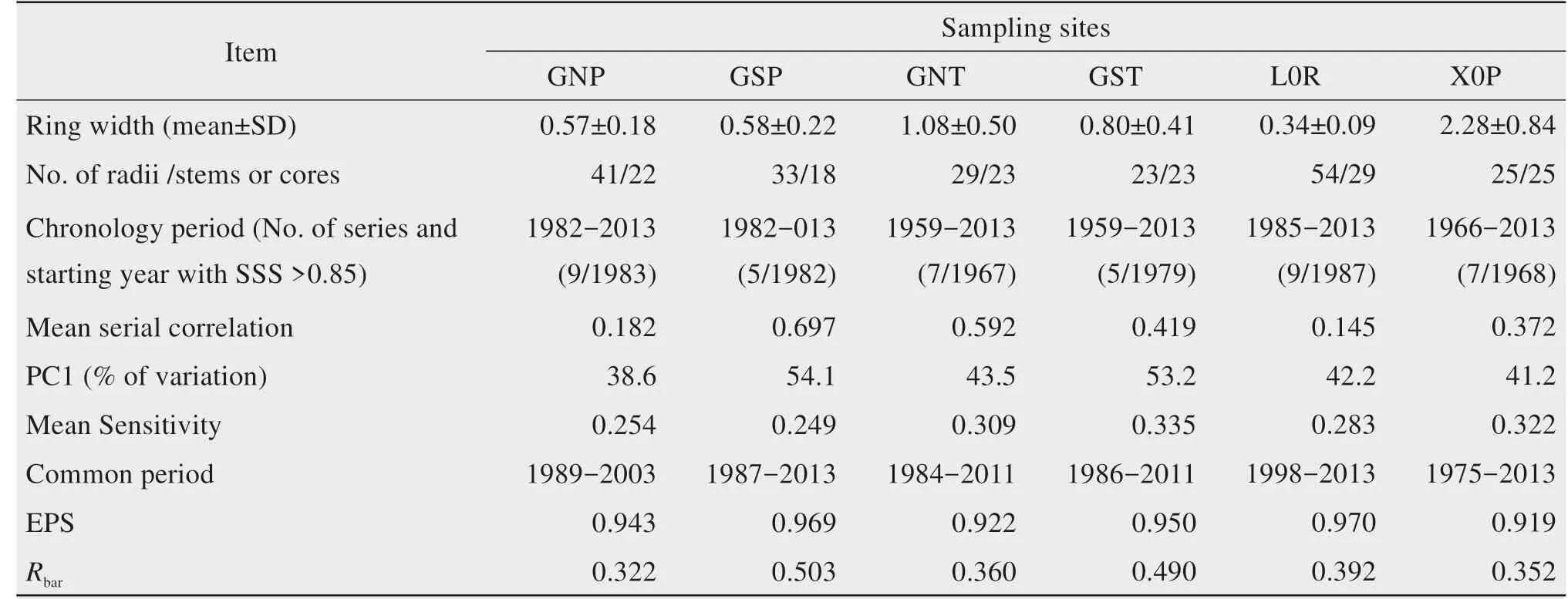
Table 2 Statistical characteristicsof the six standardized chronologiesat the sampling sites
3.3 Relationships between shrub and tree grow th and the climate,slope aspect,and management options
Figure 5 shows the resultsof the correlation analysis between tree grow th and precipitation during the growing season.At the GNPsite during the period without irrigation(GNPn;Figure 5a),grow th was significantly(P<0.05)and positively correlated with the monthly precipitation from May to July and May to August.During the irrigation period(GNPi),the ring-w idth chronology was not significantly correlated with precipitation during any month or longer period.At the GNT site(Figure 5b),the ring-w idth chronology during the period without irrigation(GNTn)was significantly(P<0.05)positively correlated with the monthly precipitation in July and from May to July.While,the ring-w idth chronology during the irrigation period(GNTi),the shrub grow th was positively but not significantly correlated with precipitation during anymonth or longer period.
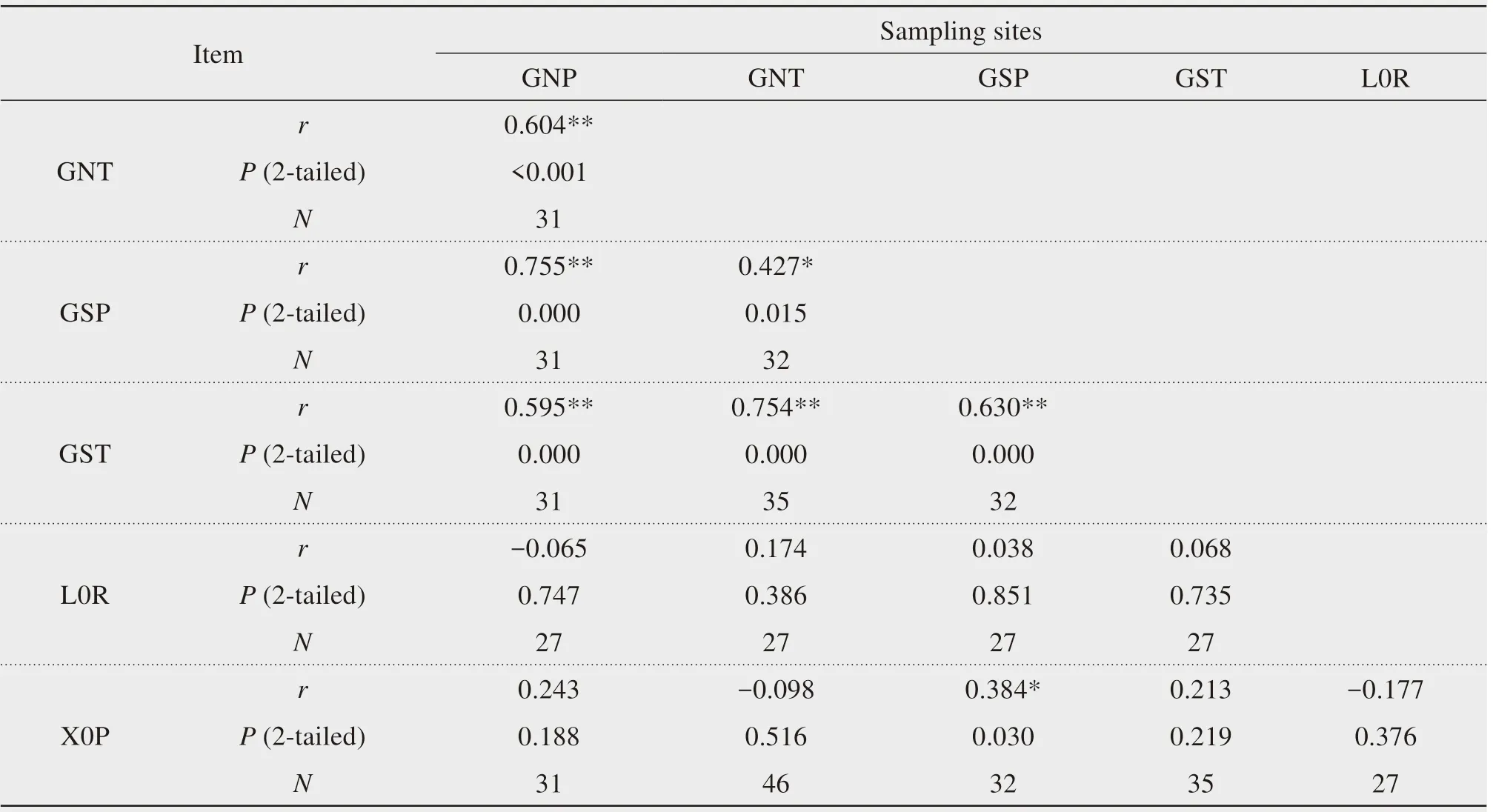
Table 3 Correlations(Pearson's r)between the chronologiesat the six sampling sites
At the GSPsite(Figure 5c),the ring-w idth chronology during the period without irrigation(GSPn)was significantly(P<0.05)positively correlated with the monthly precipitation from May to June,May to July,and May to August.During the irrigation period,the ring-w idth chronology was not significantly correlated with precipitation during anymonth or longer period.
At the GST site(Figure 5d),the ring-w idth chronology during the period without irrigation(GSTn)was significantly(P<0.05)positively correlated with themonthly precipitation from May to July and from May to August;during the period with irrigation(GSTi),no correlationwas significant.
At the X0P site(Figure 5e),the ring-w idth chronology was not significantly correlated with precipitation during anymonth or longer period during the two periods(named by X0Pi1 and X0Pi2),which was divided by the year of 1988.At the L0R site(Figure 5f),the ring-w idth chronology was significantly positively correlated with monthly precipitations in May and from May to June.
Figure 6 shows the correlations between the temperature during various months and longer periods and ring-w idth chronologies.At the GNP site(Figure 6a),with a north slope aspect,P.orientalis grow th chronology was significantly(P<0.05)negatively correlated with the mean monthly temperature in April during the irrigated period(GNPi);no other correlation was significant.At the GNT site(Figure 6b),with a north slope aspect,T.ramosissima chronology was significantly(P<0.05)negatively correlated with themeanmonthly temperature in July and Augustduring the irrigated period but not during any other month or period.
At the GSP site,with a south slope aspect,the chronology was not significantly correlated with temperature for P.orientalis(Figure 6c),but for T.ramosissima(Figure 6d),significantly negative correlations existed in July and August during the irrigated period(GSTi).No other correlationswere significant.For P.orientalis at the X 0P site(Figure 6e),there were no significant correlations between the chronologies and temperature.At the L0R site(Figure 6f),the natural dwarf shrub R.soongorica showed no significantcorrelationswith temperature.
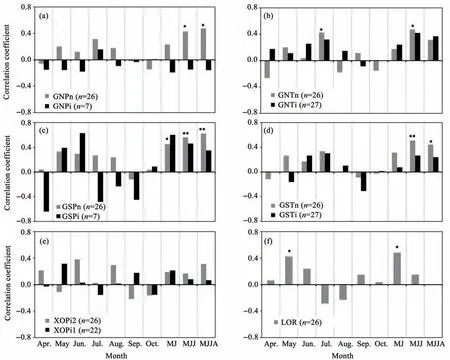
Figure 5 Correlation coefficients(Pearson's r)between the ring-w idth chronologiesat the six sampling sitesand precipitation in variousmonthsand periods(M J,May to June;M JJ,M ay,June,and July;M JJA,May,June,July,and August).Significance:*and**:P<0.05 and P<0.01,respectively.(a):GNP;(b):GNT;(c):GSP;(d):GST;(e):X0P;(f):L0P(Site code+i,with irrigation;site code+n,without irrigation;n=sample size;site code+i1 or i2:with furrow irrigation before or after the year of 1988)
4 Discussion
4.1 Influence of the regional climate on shrub and tree radial grow th
In arid areas,the recharge of soilmoisture by precipitation is crucial to sustain plant grow th(Watson and Luckman,2001,2004;Liu et al.,2013;Ren et al.,2015).In this study,radial grow th in the rain-fed artificial forest(P.orientalis and T.ramosissima)and the naturalwoody vegetation(R.soongorica)was limited by the low rainfall during the grow ing season from May to August(Figures 5a-5d and 5f).Grow th of T.ramosissima showed consistent and significant negative responses to the mean monthly air temperature duringmostof the grow ing season,both with and without irrigation and on slopeswith a north or south aspect(Figure 6).The significant correlations between the chronologiesof P.orientalis and T.ramosissima where they grew at the same sampling site(Table 3)also supported this result.Huang et al.(2006)found that the ring w idth of artificial P.orientalis forest in northern China has a significant positive response to precipitation and a significant negative response to temperature,both from June to August during the current grow ing season.Using the dendrometer,Jiang et al.(2014)found that the intra-annual stem radial grow th of P.orientalis during the main grow ing period(from m id-June to m id-September)in a sem i-arid area of northern China was influenced positively by relative hum idity and precipitation,and negatively by vapor-pressure deficitand daily airmaximum temperature,which together strongly affected moisture availability for trees.
For T.ramosissima,a phreatophyte,stem radial grow th wasmainly controlled by changes in groundwater level in a desert riparian forest(Xiao etal.,2014).A previous dendroclimatic study of R.soongorica,a dwarf shrub,on slopes with different aspects in the same area,showed a positive response tomonthly precipitation during the grow ing season sim ilar to that in the present study(Xiao etal.,2006).Additionalwater,such as that provided by irrigation or water harvesting measures(Zhang et al.,2007),w ill decrease the effect of water stress on tree grow th.Therefore,for the artificial forestduring the irrigation period,including continuous irrigation at the X0P site,tree grow th showed little or no effect of water stress,irrespective of the monthly precipitation during the grow ing season(Figure 5).This is also supported by the significantly greater annual stem radial grow th during the irrigation period for P.orientalis and T.ramosissima(Figure 3b).
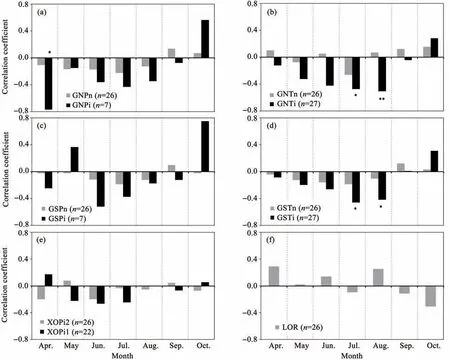
Figure 6 Correlation coefficients(Pearson's r)betweenmeanmonthly air temperature from April to Octoberof the currentyear(using data from 1954 to 2013)and ring-w idth chronologies for the six sampling sites.See Table 1 fordetailsof the site characteristics.Significance:*and**:P<0.05 and P<0.01,respectively.(a):GNP;(b):GNT;(c):GSP;(d):GST;(e):X0P;(f):L0P(Site code+i,with irrigation;site code+n,withoutirrigation;n=sample size;site code+i1 or i2:with furrow irrigation beforeorafter the yearof1988)
High air temperatures during the grow ing season increase the water consumption by increasing evapotranspiration,thereby increasing water stress and decreasing tree grow th.This explains the predom inantly negative correlations between stem radial grow th and air temperature(Figure 6).with sufficient water,plantsgrow welland produce higher biomass.However,plants with higher biomass w ill consume much more soilwater than smaller plants,possibly resulting in water stress during months with higher temperatures.A pot experiment showed that water consumption by shrubswith a sufficientwater supply was 3 to 8 times that observed under rain-fed conditions in our study area(Song et al.,2006).When the additional waterwas no longer provided,plantgrow th would decrease,aswould water consumption.Thus,P.orientalis and T.ramosissima showed a negative correlation to high temperatures(Figures 6a,6b,6c and 6d),and the responsewas significant for T.ramosissima in July and August(Figures6b and 6d).
4.2 Influence of topography on radial grow th
The topography of a site affects the solar radiation received by plants and the soil,and thereby affects evapotranspiration,which in turn affects soilmoisture and plant grow th(M iller et al.,2001;Liang et al.,2006;M?ren et al.,2015;Xiao et al.,2015).Generally,sites in the northern hem isphere with a south aspect receive more solar radiation than slopes with a north aspect,resulting in higher evapotranspiration and lower soil moisture on the south-facing sites.These differences are closely associated with differences in the natural vegetation composition and structure.In general,themore xeric and warmer south-facing slopes w ill support drought-resistant vegetation,whereasmoistand colder north-facing slopesw illsupportmoisture-loving plants(M iller etal.,2001;Liang et al.,2006;Zhong et al.,2006;M?ren et al.,2015).In our study,the stem radialgrow th of T.ramosissima differed significantly between plants grow ing on slopes with north and south aspects;itwas higher on the north-facing slope(Figure 3a;GNT and GST).This response can also be seen in the relationship between stem radial grow th and monthly precipitation on slopes with the two aspects(Figures 5b and 5d).Correlations during the periods without irrigation were generally stronger on the slopeswith a south aspect(GSPn and GSTn)than on slopeswith a north aspect(GNPn and GNTn),particularly for total precipitation during the grow ing season(starting in May and ending in June,July,or August).The correlation showed that the grow th of T.ramosissima on slopes with a south aspect ismore lim ited by soilmoisture than on slopeswith a north aspect.Thus,the stem radial grow th of T.ramosissima on the north-facing slopewasbetter than on the south-facing slope.
However,P.orientalis grow th did notdiffer significantly between the slopes with north and south aspects(GNPand GSP;Figure 3a).Thismay be related to the inherent drought resistance of this species.A study by Zhong et al.(2006)showed that the root morphology distribution pattern of this species was controlled by the different soil moisture regimes on slopeswith opposite aspects.The roots of P.orientalis on slopeswith a south aspectweremainly thin,with a diameter of less than 1mm,which increasesmoisture absorption.In contrast,the roots of plants on a slope with a north aspectweremainly thick,with a diameter greater than 3 mm.Therefore,P.orientalis can meet itswater demand by developing a different root distribution pattern on north-and south-facing slopes.
4.3 Drought resistance and adaptability of the different species
Drought resistance and adaptability are genetic traits.For example,R.soongorica is a super-xerophytic dwarf shrub,and can persist in areaswhere annual precipitation ranges from less than 50mm to 400mm(Xiao etal.,2006).In our study area,annual precipitation is 249mm,and is therefore suitable for grow th of this native shrub.The resultof a potexperiment in the same study area showed that saplings grow th of R.soongorica was no different in moderate drought stress and rain-fed conditions(Song et al.,2006).In contrast,P.orientalis developed some yellow leaves in the rain-fed treatment and T.ramosissima began to show decreased grow th and leaf w ilting in the lower water supply treatmentand partial dieback in the rainfed treatment.This shows that the drought resistance and adaptability of the indigenous species,R.soongorica,was better than that of the other species.This result agreeswith the results of our study,in which T.ramosissima grew slow ly,with only a few living branches.In addition,the ring w idth of T.ramosissima during the irrigation period was two to three times thatduring the period without irrigation(Figure 3b).
Many studies have also reported that water consumption by P.orientalis was lower than thatof some broad-leaved and coniferous trees,such as Robinia pesudoacacia and Pinus tabulaeformis(Ma et al.,2003;Wang et al.,2004;Zhang,2006;Zhang et al.,2006).
4.4 Influence of management options on stem radial grow th
Tree grow th is also affected by management options,such as the irrigation regime,planting density,and branch pruning(Zhong et al.,2006).Our results show that the stem radialgrow th of P.orientalis at the irrigated X 0P site was about 3 to 4 times that at the non-irrigated GSPand GNPsites(Table 2;Figure 3a).For T.ramosissima,stem radialgrow th during the irrigated period was significantly greater than that in the non-irrigated period on slopeswith both aspects(Figure 3b).Under irrigation,the stem radial grow th of these two species did not increase significantly with increasing precipitation(Figure 5).This shows that the additionalwater supply provided by irrigation can relieve the effects of soilmoisture stress in artificial plantations.
4.5 Implications for management of artificial forests
Pollen records in the arid western parts of the Loess Plateau since the last Glacial Maximum shows that the region's natural vegetation wasmainly herbs and drought-tolerant shrubs(Jiang et al.,2015).This is also the current status of the natural vegetation in the study area(Xiao et al.,2015).This could be one reason why R.soongorica,a native dwarf shrub,could be the preferred plant species for ecological restoration in this region.Our results show that P.orientalis can be planted on slopeswith any aspect,whereas T.ramosissima is best planted only on slopes with a north aspectunder rain-fed conditions.Both species must be assisted by artificialmeasures,such as controlling the plant density during the early stages of afforestation and other forms ofmanagement during the whole grow ing season(Xiao et al.,2015);managementmeasures therefore include branch pruning,intermediate thinning and shrub coppicing,as well as providing supplemental water by means of rainwater harvesting(e.g.,constructing counter-slopes,flat terraces,digging cone-shaped pits that increasewater infiltration into the soiland covering them with straw or plastic films)(He et al.,2002;Song et al.,2006;Zhang etal.,2014;Xiao etal.,2015).Thesemeasures w ill let the stands of woody vegetation sustain an improved long-term dynam ic water balance between consumption and wateravailability.
In addition,given the herbaceous vegetation expected to become dom inant in this region,itw ill likely provemore effective to establish a landscapemosaic that contains am ixture of trees,shrubs,and grasslands(e.g.,native steppe vegetation)to improve soil water conservation and maintain higher levels of biological diversity in the afforested land.The expensive and labor-intensive irrigation pattern used in some parts of the study area w ill only be suitable in suburban areas and local parks.Given the arid conditions in most of the Loess Plateau,planting or maintaining near-natural or natural vegetation communities seems likely to be the bestmeans of ecological restoration(Jiang etal.,2015).
5 Conclusions
In the arid western Loess Plateau of China,the radial stem grow th of R.soongorica,P.orientalis,and T.ramosissima was influenced by the regional climate,site topography(slope aspect),andmanagement options(providing supplementalwater).The precipitation during themain grow ing season was significantly positively correlated with ring grow th for all three species on slopes with both north and south aspects.A lthough water supplementation(e.g.,irrigation,rainwater harvesting)can relieve drought stress and promote tree grow th,it is likely to be too expensive and labor-intensive for w idespread use,particularly given the study area's lim ited water resources.Our results suggest that as themain afforestation species for soil and water conservation in the Loess Plateau,P.orientalis ismore suitable than T.ramosissima(a phreatophyte)under rain-fed conditions,and T.ramosissima is suitable only for slopeswith a north aspect.A landscapemosaic that combines a tree(P.orientalis)with a shrub(R.soongorica)and grassland appears likely to be the bestmeansof ecological restoration in the arid western Loess Plateau.
Acknow ledgments:
Our research was funded by the National Natural Science Foundation of China(GrantNo.41471082).
 Sciences in Cold and Arid Regions2019年3期
Sciences in Cold and Arid Regions2019年3期
- Sciences in Cold and Arid Regions的其它文章
- Vice p resident Zhang visited Cryosphere Research Station in A laska,Northwest Institute of Eco-Environment and Resource,Chinese Academ y of Sciences
- Risk prevention and controlstrategies for the severely affected areasof snow disaster in the Three Rivers Source Region(TRSR),China
- A landscapem anagementanalysis framework and its prelim inary app lication in Ejina Oasis,Northwest China
- MODISobserved snow cover variations in theAksu River Basin,Northwest China
- Assessing the im pactsof ecological-living-productive land changeson eco-environm entalquality in Xining City on Qinghai-Tibet Plateau,China
- Variation and relationship between soilm oisture and environm ental factors in the source region of the Yangtze River from 2005 to 2016