
Differences of core genes in liver fibrosis and hepatocellular carcinoma:Evidence from integrated bioinformatics and immunohistochemical analysis
2022-07-16 05:51:46YueLiShouLiYuanJingYaYinKunYangXinGangZhouWenXieQiWang
INTRODUCTION
Chronic liver infection, including chronic hepatitis B (CHB), chronic hepatitis C, alcoholic liver disease,and non-alcoholic fatty liver disease can all result in liver fibrosis. Primary liver cancer is the seventh most common cancer worldwide[1]. Hepatocellular carcinoma (HCC), which is the dominant type of liver cancer, accounts for approximately 75% of all liver cancers worldwide[2] and is the second most fatal disease in China[3]. Liver cirrhosis is an advanced stage of liver fibrosis and is characterized by limited regeneration capacity and serious complications[4]. Most HCCs develop in the background of chronic liver injury, hepatic inflammation, and liver fibrosis. Unfortunately, to date, there are still no effective treatment strategies for liver cirrhosis, and the limited number of specific biomarkers for HCC related to fibrosis further compounds the problem of its diagnosis and treatment[5,6].
He went home and from then on, he went to that store every day and bought a CD, and she wrapped it for him. He took the CD home and put it in his closet3. He was still too shy to ask her out and he really wanted to but he couldn t. His mother found out about this and told him to just ask her. So the next day, he took all his courage and went to the store as usual. He bought a CD like he did every day and once again she went to the back of the store and came back with it wrapped. He took it and when she wasn t looking, he left his phone number on the desk and ran out...
The pathogenesis of liver fibrosis and HCC is complex as the interaction of many factors may lead to their occurrence. In recent years, with the optimization of gene sequencing platforms, several differentially expressed genes (DEGs) have been identified using bioinformatics analysis[7,8]. To date, there is a huge collection of data stored in the Gene Expression Omnibus (GEO) database of gene expression that can be explored to find the relevant DEGs for a diseased condition. Chan
[9] identified DEGs between cirrhotic and non-cirrhotic livers using microarray gene analysis. Many human genes may show differential expression patterns and functions with the onset of fibrosis and/or HCC. However,the study by Chan
[9] was limited by the small sample size, which only included 24 patients with cirrhosis and 16 patients without cirrhosis. The results obtained solely from either bioinformatics or experimental approach may not elucidate relevant DEGs. Hence, integrating bioinformatics methods with experimental techniques may help us to better understand the underlying mechanisms behind fibrosis/HCC pathogenesis.
In this study, we analyzed three databases from GEO, R software packages and online tools to identify DEGs, including upregulated and downregulated genes between liver fibrosis and HCC. The molecular function, cellular component, biological process, and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways of DEGs were then assessed. We also constructed a protein-protein interaction (PPI) network using Cytoscape for further analysis. Using these methods, hub genes were identified and subjected to KEGG pathway enrichment analysis. Finally, real-time quantitative polymerase chain reaction (qPCR), Western blotting, and immunohistochemistry of the liver tissue samples from mouse model and patients were carried out to identify novel biomarkers of liver fibrosis.Our study identified fibrosis-related core genes and compared their phenotypic differences between liver fibrosis and HCC.
MATERIALS AND METHODS
Patient liver samples collection
Liver tissue samples were collected from 5 healthy controls, 40 patients with CHB (
= 28) and CHBassociated HCC (
= 12) at the Beijing Ditan Hospital, Capital Medical University, Beijing, China. The diagnosis of CHB was based on the “Guidelines on prevention and treatment of chronic hepatitis B in China”[10]. Chronic HBV infection is defined as the persistence of HBsAg in blood serum for at least 6 mo. Patients who were diagnosed for Hepatitis C viral infection, drug-induced liver disease, nonalcoholic liver disease, alcoholic liver disease, autoimmune liver disease, cholestatic liver disease, or hereditary metabolic liver disease were excluded. All samples were analyzed by a clinician and two independent pathologists with no prior knowledge of demographic and clinical data. The degree of liver inflammation and fibrosis were scored according to the METAVIR system[11], and liver samples were divided into five groups:normal control, fibrosis grade 0 (S0), fibrosis grade 1-2 (S1-2), fibrosis grade 3-4 (S3-4), and HCC group. Meanwhile, we collected the clinical data, including sex, age, HBeAg,HBV DNA, alanine transaminase, aspartate transaminase, total bilirubin, albumin, cholinesterase and alpha-fetoprotein. The study protocol conformed to the ethical guidelines of the 1975 Declaration of Helsinki, as approved by the Ethics Committee of Beijing Ditan Hospital.
Animals
Six-week-old male C57BL/6 mice were purchased from Vital River Laboratory Animal Technology Co.,Ltd, Beijing, China. All mice were housed in a specific pathogen free laboratory animal house (Institute of Zoology of Beijing, Chinese Academy of Sciences, China) at 24 ℃ with a 12 h light/dark cycle. All animal studies were approved by the Institutional Animal Care and Use Committee of the Institute of Zoology, Chinese Academy of Sciences.
Twelve male C57BL/6 mice were randomly divided into two groups:control group and carbon tetrachloride (CCl
)-treated group. To induce liver cirrhosis, CCl
(0.5 μL/g) mixed with corn oil was intraperitoneally injected into the mice three times per week for 12 wk. The control group was injected with an equal volume of corn oil. CCl
was purchased from Sigma-Aldrich (St. Louis, MO, United States).
Prior to Holly s birthday, Susan had been a regular visitor in our home. On several occasions, she rode the bus home with Holly and was one of the few friends ever permitted to stay over on a school night. The girls did their homework together and went to bed at a reasonable hour.
The island s owner, Vlado Juresko said, We think it is the most perfect heart-shaped island in the world. Nobody lives there so if lovers really do want to spend time alone it s the perfect desert island.
Histological and immunohistochemical analysis
For hematoxylin and eosin (HE) staining, Masson’s trichrome staining, and immunohistochemical analysis, liver tissues collected from mouse model or patients were fixed with 4% paraformaldehyde solution and embedded in paraffin. For histological analysis, 5 μm thick sections were stained and observed under 10 × or 20 × objective lens. Masson’s trichrome kit (G1281, Solarbio, Beijing, China) was used according to the manufacturer’s instructions.
Suddenly, out of breath, she snatched the tabor from Abdallah with her left hand, and, holding the dagger in her right hand, held out the tabor to her master
For immunohistochemical staining, 5% bovine serum albumin in 0.1% TritonX-100 tris-buffered saline was used as the blocking solution. The samples were incubated overnight at 4 ℃ with antidecorin (DCN) (ab277636, Abcam), anti-dermatopontin (DPT) (10537-1-AP, Proteintech), and anti-SRYbox transcription factor 9 (SOX9) (ab185966; Abcam) antibodies. After incubation with a peroxidaseconjugated secondary antibody, the signal was visualized using a diaminobenzidine peroxidase substrate kit. The collagen area or positive area of immunohistochemical staining was quantified using ImageJ 1.52a software.
Real-time qPCR
Total RNA from liver tissues was isolated using TRIzolTM reagent (Thermo Fisher Scientific, MA,United States). Isolated RNA was reverse-transcribed into complementary DNA (cDNA) using a highcapacity cDNA reverse transcription kit (Promega, WI, United States). The relative expression of genes was detected by real-time fluorescence qPCR system (Light Cycler 480, Roche, Sweden) with SYBR green master mix (Promega, WI, United States). The primer sequences used in this study were listed in Supplementary Table 1. Statistical significance between the control and CCl
-treated groups was defined at
< 0.05.
Western blotting
Bioinformatics has screened out many differentially expressed genes related to liver fibrosis; however, it is unknown whether these genes are different in animal and human liver fibrosis tissues, especially among the different fibrotic degrees. Therefore, we should carefully analyze the research results of bioinformatics.
Microarray and gene expression analysis
The RNA expression data of human cirrhotic and healthy livers were collected from the GEO database (http://www.ncbi.nlm.nih.gov/geo). The GSE14323, GSE36411, and GSE89377 datasets contained gene expression data collected from 41, 21, and 12 cirrhotic and 19, 21, and 13 healthy liver tissues,respectively. The GEOquery R software package was used to download the GEO data and platform information. Then, the gene ID conversion was performed, and the maximum value of genes with the same name was selected. Ggplot2 package was used to plot the boxplot and density of the expression levels for each sample. Ggfortify package was used to perform the principal component analysis (PCA).DEGs between the cirrhosis and healthy liver tissue groups were identified using limma package by limiting the value of adjustment:
-value (adjust.
< 0.05) and the absolute value of logFC (|logFC| >1.2 or 0.6). A volcano map was generated using the ggplot2 package, and Venn diagram online tool (http://bioinformatics.psb.ugent.be/webtools/Venn/) was used to draw a Venn map. The DEGs were verified using the ONCOMINE server (https://www.Oncomine.org/resource/), which is an online available microarray database[12].
Gene ontology and pathway enrichment analysis
WebGestalt online tool (http://www.webgestalt.org) was used to identify DEGs enriched in biological processes, molecular functions, cellular component-related pathways, and KEGG pathways. The
value of less than 0.05 was considered statistically significant.
PPI network construction and identification of hub genes
STRING online database (https://string-db.org/; version 11.5) was used to build the PPI network[13].The DEGs were submitted to the STRING database to construct the PPI network. Cytoscape (version 3.7.2) was used to draw the PPI network of DEGs, and the cytoHubba plugin was used to identify hub genes[14].
Gene expression level in HCC and survival analysis
Gene expression profiling interactive analysis (GEPIA) online server (http://gepia.cancer-pku.cn/) was used to analyze the RNA sequencing expression data of tumors and healthy samples from the cancer genome atlas and genotype tissue expression projects[15]. We used this server to check whether the identified hub genes were differentially expressed in HCC tissues. Overall 5-year survival rates according to gene expression were obtained using the Kaplan-Meier Plotter (http://kmplot.com/analysis/index.php?p=service&cancer=liver_rnaseq)[16].
Statistical analyses
GraphPad Prism 6.0 (GraphPad Software Inc. La Jolla, CA, United States) was used for statistical analysis. Data were presented as mean ± SE or SD (for normally distributed data) or median with interquartile range (for non-normally distributed data). Statistically significant differences were determined using a two-tailed Student’s
-test or analysis of variance (ANOVA). Statistical significance was set and marked as
< 0.05,
< 0.01,
< 0.001, and
< 0.0001. Replicates are indicated in the figure legends, and (
) represents the number of experimental replicates.
RESULTS
Identification of DEGs
We screened and identified 10 hub genes related to fibrosis. The expression levels of DCN, DPT, and SOX were positively correlated with the degree of liver fibrosis but showed different correlations with the survival rate of patients with HCC.
Gene Ontology and KEGG pathway analysis of DEGs
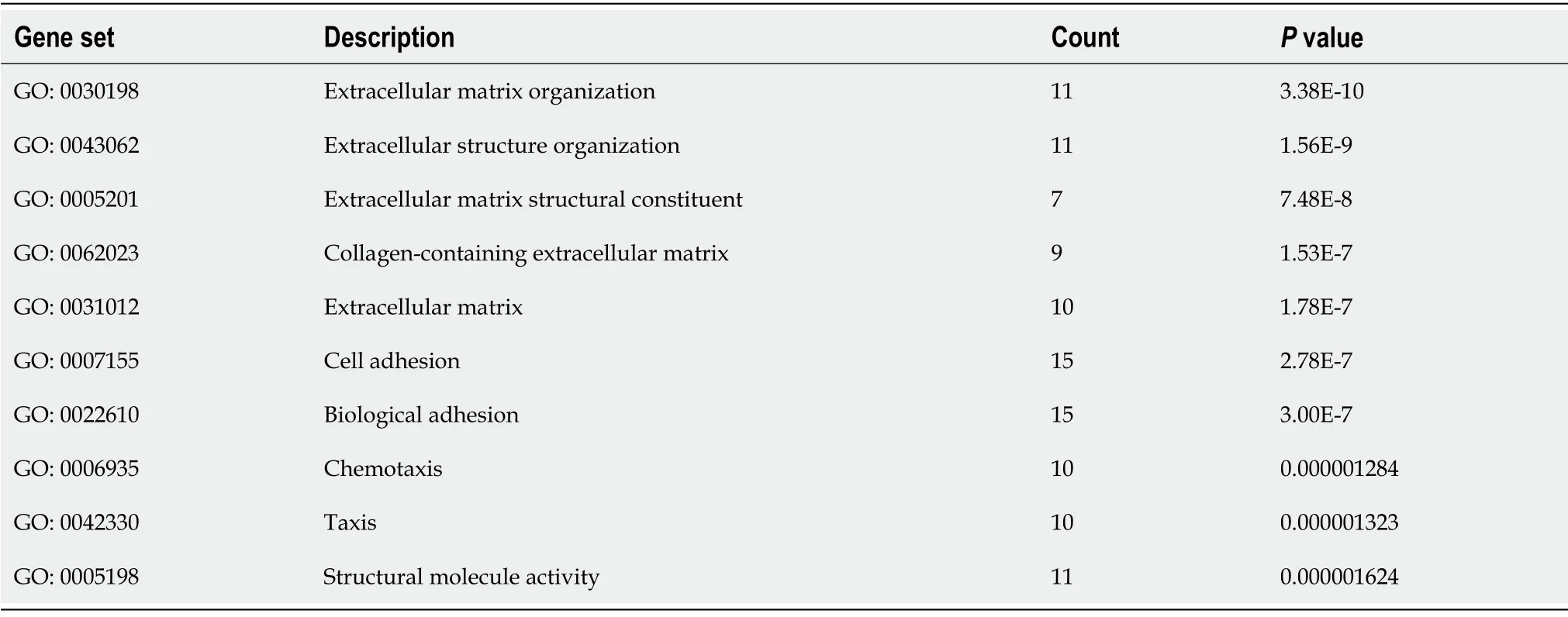
To understand the function of identified DEGs, we carried out Gene Ontology (GO) and KEGG pathways analyses using Webgestalt online server. The GO analysis results were listed according to
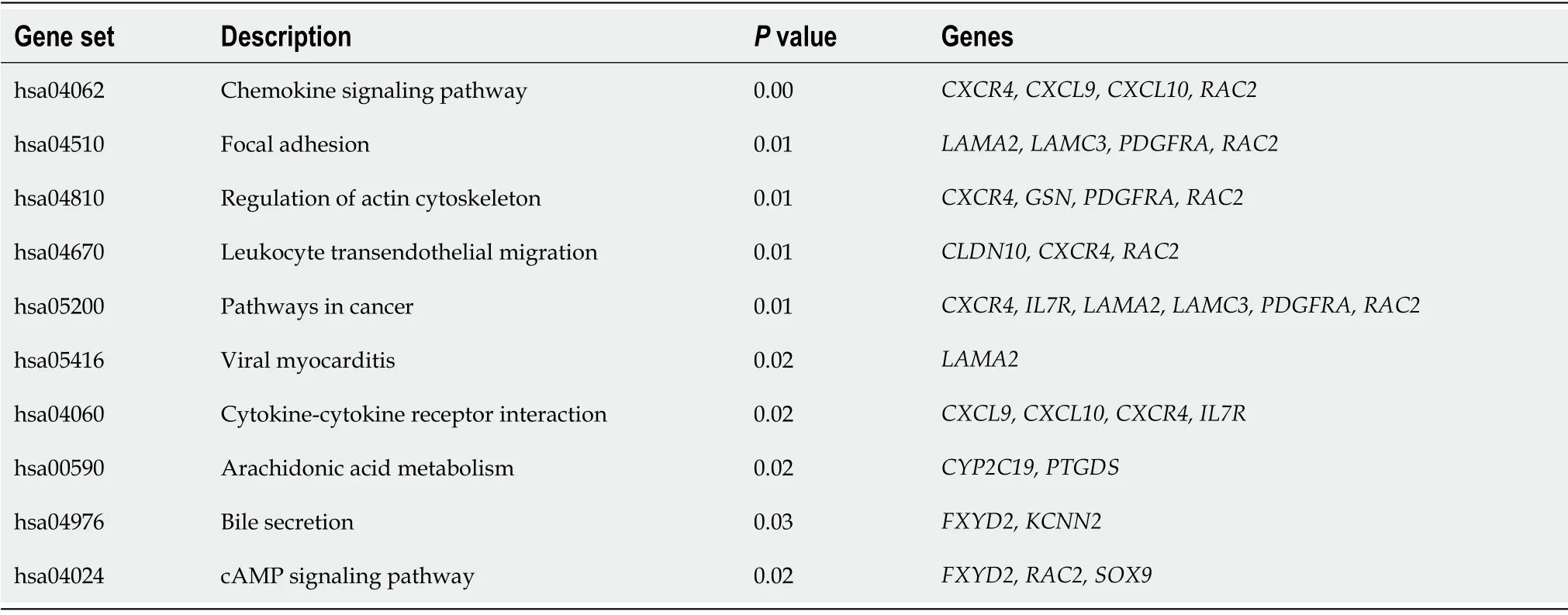
values (Table 2), which showed that DEGs were significantly enriched in extracellular matrix organization (GO:0030198), extracellular matrix structural constituent (GO:0005201), collagencontaining extracellular matrix (GO:0062023), extracellular matrix (GO:0031012), cell adhesion (GO:0007155), biological adhesion (GO:0022610), and taxis (GO:0042330). KEGG pathway analysis showedthat the DEGs involved in the chemokine signaling pathway, focal adhesion, regulation of actin cytoskeleton, leukocyte transendothelial migration, pathways in cancer, and cytokine–cytokine receptor interaction, and arachidonic acid metabolism were highly enriched in patients with liver cirrhosis(Table 3).

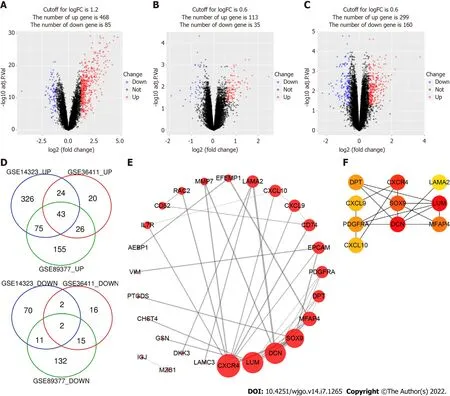
PPI network construction and identification of hub genes
PPI network analysis aids in studying the molecular mechanisms of the disease pathogenesis. Using String v11 and Cytoscape software, we constructed a PPI network with 26 nodes and 36 edges(Figure 1E). These genes were upregulated in liver cirrhosis. The top 10 hub genes were identified using the CytoHubba plugin of Cytoscape, which included C-X-C motif chemokine ligand 9 (
),
C-X-C motif chemokine receptor 4 (
),
,
laminin subunit alpha 2 (
),lumican (
), microfibril associated protein 4 (
), platelet-derived growth factor receptor alpha(
), and
(Figure 1F).
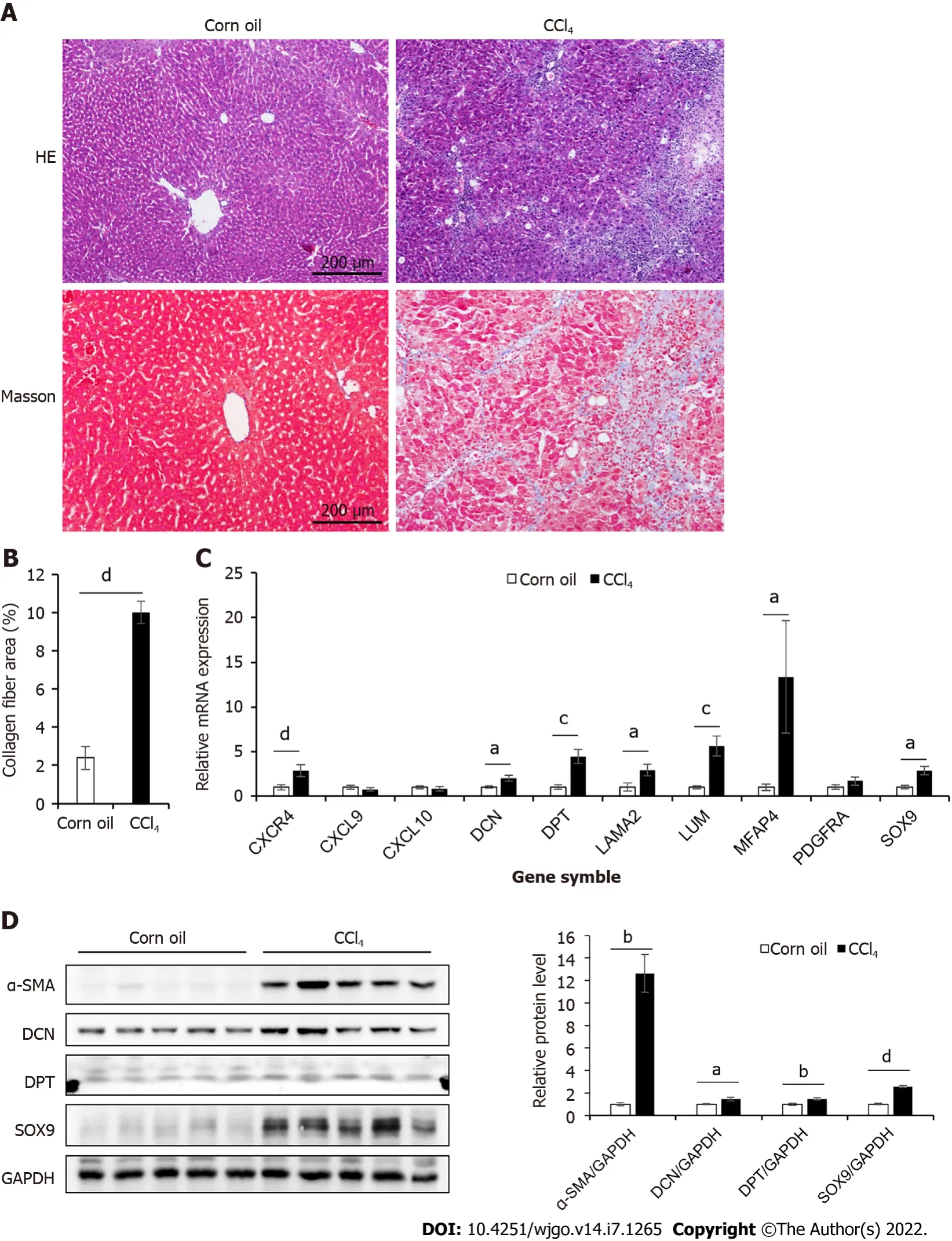
Expression of hub genes in the liver tissue of CCl4-induced cirrhosis mice
To study the role of the 10 hub genes in liver cirrhosis, we generated a CCl
-induced liver cirrhosis mouse model (Figure 2A–B). The relative mRNA levels of these genes were shown in Figure 2C.Compared to those in the control group, seven genes (
and
) were significantly upregulated in the liver cirrhosis mice, while the expression levels of
, and
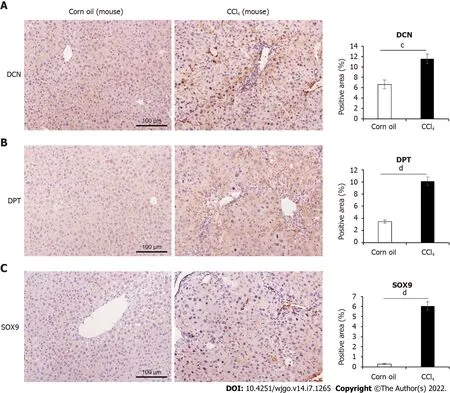
were not significantly different between the two groups. For further validation, we performed Western blotting and immunohistochemical analysis (Figure 2D, Figure 3).These results confirmed that the protein levels of DCN, DPT, and SOX9 were significantly upregulated in the liver tissue of cirrhotic mice.
DCN, DPT and SOX9 expression and liver fibrosis progression in patients
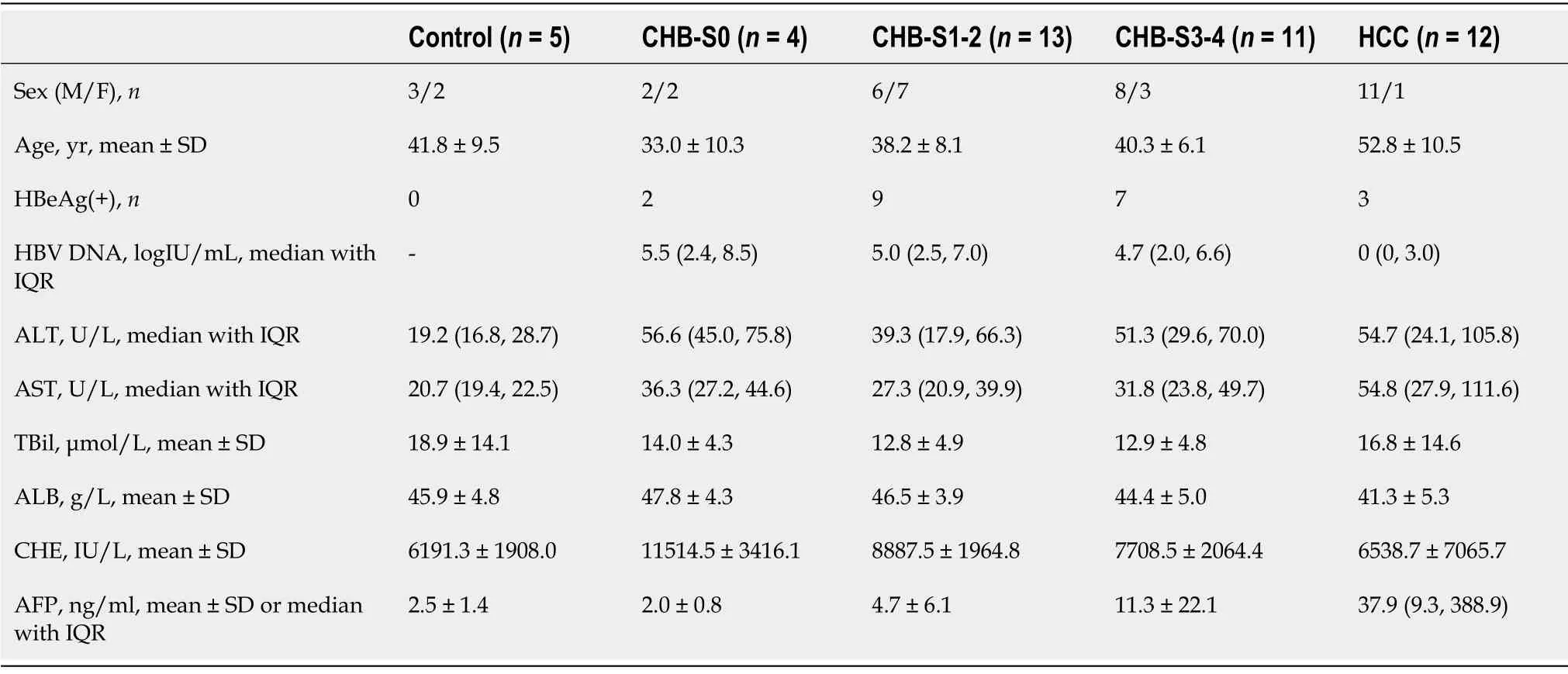
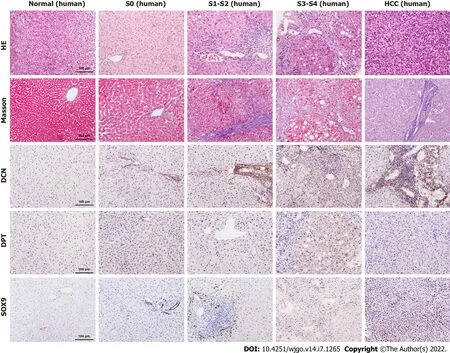
To further explore the relationship between DCN, DPT, and SOX9 protein expression and progression of liver cirrhosis, we collected liver biopsy tissue samples from 5 healthy controls, 28 patients with CHB,and 12 patients with CHB-associated HCC. All liver tissues were divided into 5 groups (normal, S0, S1-2, S3-4, and HCC) according to METAVIR system. The clinical profile of the patients enrolled in the study is summarized in Table 4. The results showed that males were the majority in S3-4 and HCC groups. The age, TBil, ALB, and CHE values in all groups and the AFP value in non-HCC group were in normal distribution, and the median AFP value in the HCC group was higher than the upper limit of normal value. In patients with CHB, HBV DNA was detected as positive, and most ALT and AST levels were elevated, which was consistent with the inflammatory activity of the liver. Most patients with HCC were detected negative for HBV DNA, which is related to antiviral treatment. Liver samples from patients with fibrosis showed increased collagen deposition, inflammatory cell infiltration, or atypical cells viewed with HE and Masson’s trichrome staining (Figure 4).
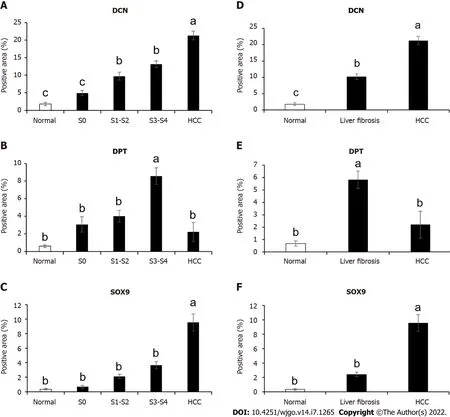
Immunohistochemical results showed that the expression levels of DCN and SOX9 increased with the aggravation of liver fibrosis (Figure 4, Figure 5A, Figure 5C) and were significantly higher in the HCC group than those in healthy controls and fibrotic groups (Figure 4, Figure 5D, and Figure 5F). Further,compared with healthy controls, DPT expression was significantly increased in patients with liver fibrosis, particularly in the S3-S4 group but extremely reduced in patients with HCC (Figure 5B,Figure 5E). In addition, we found that DCN was mostly expressed in the portal vein region, which was highly consistent with the distribution of collagen fibers, while SOX9 and DPT were mostly expressed in hepatocytes and several other types of cells (Figure 4).
DCN, DPT, and SOX9 expression and survival rate of patients with HCC
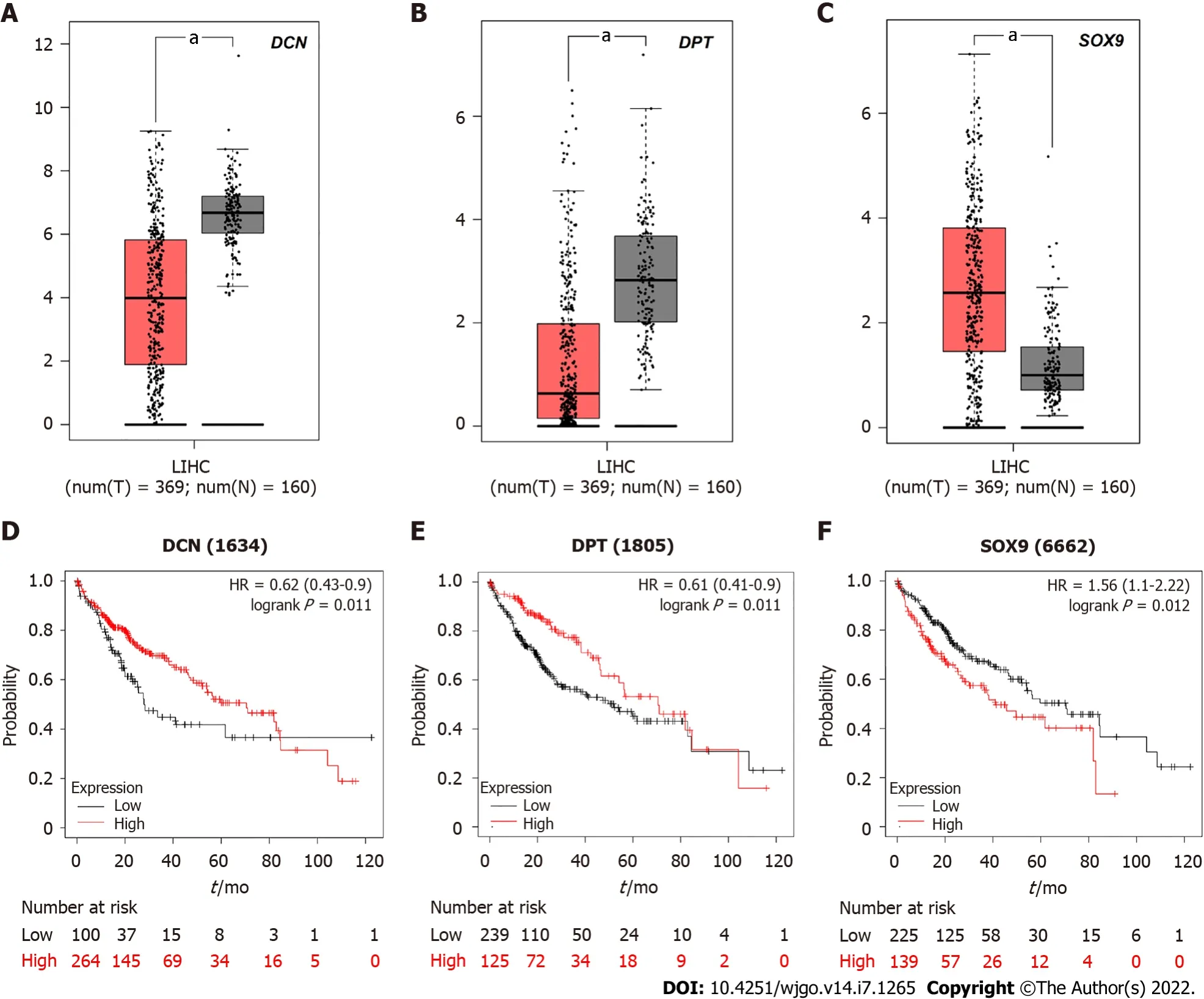
Due to the differential expression pattern and distribution, we decided to analyze the correlation between DCN, DPT, and SOX9 expression levels and the survival rate of patients with HCC. The GEPIA server was used to detect the mRNA expression levels in tissues of patients 369 liver hepatocellular carcinoma (LIHC) and 160 normal tissues (Figure 6A-C). The results showed that SOX9 was significantly upregulated whereas DCN and DPT were significantly downregulated in patients with LIHC.
Kaplan-Meier curves, depicting samples from these 369 patients with HCC, showed that the 5-year survival rate of patients with high expression of SOX 9 was significantly reduced, while that of patients with high expression of DCN or DPT was increased (Figure 6D-F).
DISCUSSION
To identify liver fibrosis-related core genes, we observed and compared the differential expression pattern of core genes in patients with liver fibrosis and HCC.
GEO is an international public resource bank for high-throughput microarray and next-generation sequencing of functional genome datasets submitted by research groups. In this study, we identified 45 DEGs from GSE14323, GSE36411, and GSE89377 datasets in the GEO database for an in-depth analysis of their biological function. Most of these DEGs have been associated with liver diseases.
and
are involved in HCV or HBV infection[18-23].
and
are overexpressed in non-alcoholic steatohepatitis (NASH)[24-26].
and
are associated with cholestasis[27-31].
and
are upregulated in patients with HCC[32-38]. The combined data-mining of three datasets from different sources yielded 45 DEGs, further indicating that the GEO database is indeed a useful resource for understanding the mechanism of liver diseases and using the GEO database can increase the efficiency of published resources.
We screened GEO databases and obtained 45 DEGs and 10 hub genes (particularly
and
) in cirrhotic liver tissues. Upregulated expression of DCN, DPT, and SOX9 was all positively correlated with the degree of fibrosis, but there may be differences between their correlation with the 5-year survival rate of HCC patients.
Li Y and Yuan SL contributed equally to this work; Wang Q and Xie W designed the research and were co-corresponding authors; Yuan SL, Li Y, Yin JY, Yang K and Zhou XG performed the experiments; Yuan SL and Li Y analyzed the data; Yuan SL, Li Y and Wang Q wrote the manuscript; Yuan SL provided vital reagents and analytical tools; all authors read and approved the final manuscript.
As a result, I started to feel like Snow White s wicked(,) stepmother, plotting against the princess as I willed her to succumb9 to my culinary magic. Chuck, who at first ate anything and everything, developed critical tendencies. He soon took up the hue10 and cry() of Mama doesn t make it that way.
In other variants, the eldest daughter is covered with tar27 or pitch that cannot be removed, given a box of serpents, riddled64 with leprosy, or cursed with eternal ugliness.Return to place in story.
The
gene encodes a member of the small leucine-rich proteoglycan family of proteins, which can act as a tumor repressor in a variety of cancers[39]. DCN is a regulator of matrix assembly and not only targets transforming growth factor-beta 1 (TGF-β1) but is also involved in the maturation of collagen fibrils[40,41]. The enhanced deposition of DCN reflects the stimulatory effect of overproduction of TGFβ1[41]. Dudás
[42] indicated that high amounts of TGF-β1 colocalize with DCN within the fibrotic areas of the liver using a cohort of liver pathologies, including chronic hepatitis, fibrosis, and cirrhosis,which is consistent with our results. However, contrary to our results, Shang
[43] found DCN mRNA expression to be downregulated and not upregulated in patients with HCC
gene expression profile analyses. Although GEPIA analysis showed that the mRNA expression level of DCN in ILHC was lower than that normal tissues, the heterogeneity within each group, especially the ILHC group,was very different. In addition, our results were based on immunohistochemical analysis, different from the expression profile of the whole liver tissue used by Shang
[43]. From the distribution of immunohistochemical sections, we concluded that almost all the increase in DCN expression was localized in the collagen-intensive area of the portal region and not in the hepatic lobule, which indicates that the upregulated DCN significantly represents an increase in matrix assembly. These may partly explain why the increased DCN is not associated with the lower 5-year survival rate of HCC patients, and why the expression pattern of DCN expression analyzed by GEPIA server was different from our study. Considering these inconsistent results, the expression, location and function of DCN deserves further study.
Like his brothers, he made the long journey to the city of Quimus the son of Timus; like them he saw the citadel, but he saw there the heads of Tahmasp and Qamas
DPT is a downstream target of the vitamin D receptor. Fu
[44] reported that mRNA expression of DPT was significantly downregulated in HCC, while its protein was weakly expressed in tumorous tissues compared to that in non-tumorous tissues. However, Lefebvre
[45] suggested that DPT is upregulated in active NASH and fibrosis, and it is necessary for collagen deposition in profibrotic conditions. Our results also confirmed that the expression of DPT increased with the aggravation of liver fibrosis. Interestingly, a previous study showed that DPT interacts with DCN, which influences collagen fibrillogenesis and increases TGF-β1 signaling[46]. Our study identified these two molecules in a combined screening, suggesting that both DPT and DCN play an important role in the occurrence and development of liver fibrosis and HCC. However, the interactions between them needs to be further studied.
During tumorigenesis, SOX9 is upregulated in various tumors and plays an essential role in tumor progression as an oncogene[29], which regulates cellular proliferation, senescence, and self-renewal and is highly expressed in liver cancer stem cells[47]. In addition, SOX9 was the earliest marker expressed by biliary precursors[48]. It has been confirmed as a transcription factor that regulates bile duct development and contributes to liver regeneration and fibrosis[47]. In this study, we confirmed that SOX9 was positively correlated with the degree of fibrosis, and the high expression of SOX9 indicated a decline in the 5-year survival rate of patients with HCC, which is consistent with the results of other studies. In addition, our results showed that SOX9 was enriched and expressed in bile duct cells in mouse and human fibrotic livers, and the expression levels of SOX9 in hepatocytes were also increased significantly in patients with HCC. This observation can be explained by the fact that SOX9 mediates the transdifferentiation of hepatocytes into bile duct epithelial cells[47]. However, the detailed molecular mechanism of SOX9 overexpression in hepatocytes requires further elucidation.
Our current study has some limitations. First, we only analyzed the transcriptome, and many studies have shown that epigenetic modifications and non-coding RNAs also play an important role in the progression of liver diseases[33,49]. Secondly, the sample size in terms of number of patients was small,and immunohistochemistry was a semi-quantitative method. In addition, although our study had identified the signal transduction pathway involved in liver cirrhosis and HCC, it lacked in-depth analysis on the mechanism of action of these molecules, which needed to be further studied using
studies or knock-out mice.
CONCLUSION
There we sat, side by side, watching the sun go down behind the old Texaco service station across the busy street. A street I had been allowed to cross by myself. Grampy stretched his long legs down over two stairs. I dangled22 mine, a bit closer to the first step this time, I m sure.
ARTICLE HIGHLIGHTS
Research background
Liver fibrosis and hepatocellular carcinoma (HCC) are common adverse consequences of chronic liver injury. Establishing more effective biomarkers is important for understanding the pathogenesis,occurrence, development mechanisms of liver fibrosis and HCC, as well as to identify new diagnostic and therapeutic tools.
Long, long ago, in the days when fairies, witches, giants and ogres still visited the earth, there lived a king who reigned1 over a great and beautiful country
The largest of them extended for several miles; they were all lighted up by the vivid light of the aurora1, and they were so large and empty, so icy cold and glittering! There were no amusements here, not even a little bear’s ball, when the storm might have been the music, and the bears could have danced on their hind2 legs, and shown their good manners
Research motivation
Total protein from liver tissues was extracted using radioimmunoprecipitation assay buffer. The protein concentration of the samples was measured by bicinchoninic acid assay. The same concentration of protein was loaded to 10% sodium dodecyl sulfate polyacrylamide gel and then transferred to a polyvinylidene fluoride membrane. The membranes were incubated overnight at 4℃ in anti-α-SMA(ab5694, Abcam), anti-DCN (ab277636, Abcam), anti-DPT (10537-1-AP, Proteintech), and anti-SOX9(ab185966, Abcam) antibodies. All signals were visualized by density scanning (Image Quant TL7.0; GE Healthcare Biosciences, Uppsala, Sweden). The intensity of the bands was analyzed using ImageJ 1.52a software.
Research objectives
Fibrosis is a common pathological symptom of severe liver damage caused by various chronic liver diseases. The most common primary liver cancer, HCC, occurs most often in people inflicted with chronic liver diseases[17]. Understanding the molecular mechanisms underlying cirrhosis can help in the development of effective treatments. Although bioinformatics tools can be used to study the relationship between gene function, liver fibrosis, and HCC, it is necessary to comprehensively analyze gene expression levels and distribution in the context of disease pathology
experiments.
Research methods
In this study, we analyzed the expression pattern of hub genes of fibrosis and HCC. Bioinformatics analyses, quantitative polymerase chain reaction, Western blot, and immunohistochemistry of liver tissues from mouse model and patients were performed to identify novel biomarkers of liver fibrosis and HCC.
Research results
Ten hub genes (CXCL9, CXCL10, CXCR4, DCN, DPT, LAMA2, LUM, MFAP4, PDGFRA, and SOX9)associated with cirrhosis were screened from GSE14323, GSE36411, and GSE89377 datasets. DCN, DPT,and SOX9 were highly expressed in the CCl4-induced mouse model of liver cirrhosis and fibrotic patient liver samples, and their expression levels were associated with the degree of fibrosis. In patients with HCC, SOX9 was upregulated, while DCN and DPT were downregulated. However, the 5-year survival rate of HCC patients with high SOX 9 expression was significantly reduced, which is different from DPT or DCN.
Research conclusions
We analyzed three gene expression datasets, GSE14323, GSE36411, and GSE89377, which included data from 74 cirrhotic and 53 healthy liver tissue samples in total. The analysis of processed sample data showed that the gene expression levels in different samples were primarily the same (Supplementary Figure 1). PCA showed that cirrhotic and healthy groups constituted individual clusters(Supplementary Figure 2). Using limma package, we extracted 553, 148, and 459 DEGs from GSE14323,GSE36411, and GSE89377 datasets, respectively (Figure 1A-C). Using Venn diagram online tool, we identified 45 DEGs (43 upregulated genes and 2 downregulated genes) associated with cirrhosis from the three datasets (Figure 1D, Table 1).
Research perspectives
The integrated approach of bioinformatics and molecular biology is more efficient to research multifactorial diseases, such as liver fibrosis and liver cancer. Future studies on the differences on DCN, DPT,and SOX9 expression may help in the better understanding of the mechanisms involved in the development of liver fibrosis and HCC.
Next, for detailed characterization of hub gene functions, we selected three representative genes (
and
), since their expression was consistently found to be associated with liver cirrhosis. We established a CCl
-induced mice model and verified their mRNA and protein levels using qPCR, Western blotting, and immunohistochemistry to confirm the expression and main pathophysiological functions. The results confirmed that DCN, DPT, and SOX9 were significantly overex-pressed in human and mouse fibrotic liver; however, there were differences in the relationship between their expression levels and the survival rate of patients with HCC.
the Beijing Natural Science Foundation, No. 7222097; Beijing Hospitals Authority the Digestive Medical Coordinated Development Center, No. XXZ0401; National Natural Science Foundation of China, No. 82000555 and No. 81900547; and Beijing Municipal Science and Technology Commission, No. D171100003117005.
The study protocol conforms to the ethical guidelines of the 1975 Declaration of Helsinki as reflected in a priori approval by the Ethics Committee of Beijing Ditan Hospital No. 2021-034-01.
Patients were not required to give informed consent to the study because the analysis used anonymous clinical data and liver tissue samples.
All the authors report no relevant conflicts of interest for this article.
Publicly available datasets were analyzed in this study, which can be found here:GSE14323,GSE36411 and GSE89377. Technical appendix, statistical code, and data set available from the corresponding author at wangqidl04@ccmu.edu.cn. No additional data are available.
Arnoldus survived his wounds and mended slowly. When he could finally leave his bed, he sat by the window staring out into the garden. He so regretted that he hadn t given the bulbs to his family sooner. The war was over and spring was coming, but life remained very hard. Many houses had been bombed. There was little food and jobs were few.
This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is noncommercial. See:https://creativecommons.org/Licenses/by-nc/4.0/
Now it happened that three tailors had met together, and the two elder thought, that after having successfully put in so many fine and strong stitches with never a wrong one amongst them, they were certain to do the right thing here too
China
11. Heard: Some critics have considered Hansel and Gretel to be a subversive65 tale, encouraging children to eavesdrop66 on their parents, trespass67, commit murder, and steal property. The children are not ideal role models in the conservative sense, but one can credit them for being survivors68 in a harsh world. If they had not done these things, they would most likely be dead.Return to place in story.
Yue Li 0000-0002-7412-7632; Shou-Li Yuan 0000-0002-4733-9891; Jing-Ya Yin 0000-0002-9707-2363; Kun Yang 0000-0003-0688-4962; Xin-Gang Zhou 0000-0002-4211-3017; Wen Xie 0000-0002-7314-8175; Qi Wang 0000-0002-0269-1568.
Beijing Medical Association Hepatology Branch.
Gong ZM
A
Li X
1 Bosetti C, Turati F, La Vecchia C. Hepatocellular carcinoma epidemiology.
2014; 28:753-770 [PMID:25260306 DOI:10.1016/j.bpg.2014.08.007]
2 Harris PS, Hansen RM, Gray ME, Massoud OI, McGuire BM, Shoreibah MG. Hepatocellular carcinoma surveillance:An evidence-based approach.
2019; 25:1550-1559 [PMID:30983815 DOI:10.3748/wjg.v25.i13.1550]
3 Chen Z, Xie H, Hu M, Huang T, Hu Y, Sang N, Zhao Y. Recent progress in treatment of hepatocellular carcinoma.
2020; 10:2993-3036 [PMID:33042631]
4 Chen W, Zheng R, Baade PD, Zhang S, Zeng H, Bray F, Jemal A, Yu XQ, He J. Cancer statistics in China, 2015.
2016; 66:115-132 [PMID:26808342 DOI:10.3322/caac.21338]
5 Zhou WC, Zhang QB, Qiao L. Pathogenesis of liver cirrhosis.
2014; 20:7312-7324 [PMID:24966602 DOI:10.3748/wjg.v20.i23.7312]
6 Thomson MJ, Lok AS, Tapper EB. Optimizing medication management for patients with cirrhosis:Evidence-based strategies and their outcomes.
2018; 38:1882-1890 [PMID:29845749 DOI:10.1111/liv.13892]
7 Huang X, Li Y, Guo X, Zhu Z, Kong X, Yu F, Wang Q. Identification of differentially expressed genes and signaling pathways in chronic obstructive pulmonary disease
bioinformatic analysis.
2019; 9:1880-1899 [PMID:31419078 DOI:10.1002/2211-5463.12719]
8 Jiang CH, Yuan X, Li JF, Xie YF, Zhang AZ, Wang XL, Yang L, Liu CX, Liang WH, Pang LJ, Zou H, Cui XB, Shen XH,Qi Y, Jiang JF, Gu WY, Li F, Hu JM. Bioinformatics-based screening of key genes for transformation of liver cirrhosis to hepatocellular carcinoma.
2020; 18:40 [PMID:32000807 DOI:10.1186/s12967-020-02229-8]
9 Chan KM, Wu TH, Wu TJ, Chou HS, Yu MC, Lee WC. Bioinformatics microarray analysis and identification of gene expression profiles associated with cirrhotic liver.
2016; 32:165-176 [PMID:27185598 DOI:10.1016/j.kjms.2016.03.008]
10 Chinese Society of Infectious Diseases; Chinese Medical Association; Chinese Society of Hepatology, Chinese Medical Association. [The guidelines of prevention and treatment for chronic hepatitis B (2019 version)].
2019; 27:938-961 [PMID:31941257 DOI:10.3760/cma.j.issn.1007-3418.2019.12.007]
11 Bedossa P, Poynard T. An algorithm for the grading of activity in chronic hepatitis C. The METAVIR Cooperative Study Group.
1996; 24:289-293 [PMID:8690394 DOI:10.1002/hep.510240201]
12 Rhodes DR, Kalyana-Sundaram S, Mahavisno V, Varambally R, Yu J, Briggs BB, Barrette TR, Anstet MJ, Kincead-Beal C, Kulkarni P, Varambally S, Ghosh D, Chinnaiyan AM. Oncomine 3.0:genes, pathways, and networks in a collection of 18,000 cancer gene expression profiles.
2007; 9:166-180 [PMID:17356713 DOI:10.1593/neo.07112]
13 Szklarczyk D, Gable AL, Lyon D, Junge A, Wyder S, Huerta-Cepas J, Simonovic M, Doncheva NT, Morris JH, Bork P,Jensen LJ, Mering CV. STRING v11:protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets.
2019; 47:D607-D613 [PMID:30476243 DOI:10.1093/nar/gky1131]
14 Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T. Cytoscape:a software environment for integrated models of biomolecular interaction networks.
2003; 13:2498-2504[PMID:14597658 DOI:10.1101/gr.1239303]
15 Tang Z, Li C, Kang B, Gao G, Zhang Z. GEPIA:a web server for cancer and normal gene expression profiling and interactive analyses.
2017; 45:W98-W102 [PMID:28407145 DOI:10.1093/nar/gkx247]
16 Menyhárt O, Nagy á, Gy?rffy B. Determining consistent prognostic biomarkers of overall survival and vascular invasion in hepatocellular carcinoma.
2018; 5:181006 [PMID:30662724 DOI:10.1098/rsos.181006]
17 Müller M, Bird TG, Nault JC. The landscape of gene mutations in cirrhosis and hepatocellular carcinoma.
2020;72:990-1002 [PMID:32044402 DOI:10.1016/j.jhep.2020.01.019]
18 Marrocco C, Rinalducci S, Mohamadkhani A, D'Amici GM, Zolla L. Plasma gelsolin protein:a candidate biomarker for hepatitis B-associated liver cirrhosis identified by proteomic approach.
2010; 8 Suppl 3:s105-s112 [PMID:20606740 DOI:10.2450/2010.017S]
19 Guzmán-Fulgencio M, Berenguer J, Jiménez-Sousa MA, Pineda-Tenor D, Aldámiz-Echevarria T, García-Broncano P,Carrero A, García-álvarez M, Tejerina F, Diez C, Vazquez-Morón S, Resino S. Association between IL7R polymorphisms and severe liver disease in HIV/HCV coinfected patients:a cross-sectional study.
2015; 13:206 [PMID:26123260 DOI:10.1186/s12967-015-0577-y]
20 Xiang Y, Tang JJ, Tao W, Cao X, Song BL, Zhong J. Identification of Cholesterol 25-Hydroxylase as a Novel Host Restriction Factor and a Part of the Primary Innate Immune Responses against Hepatitis C Virus Infection.
2015;89:6805-6816 [PMID:25903345 DOI:10.1128/JVI.00587-15]
21 Shrivastava S, Meissner EG, Funk E, Poonia S, Shokeen V, Thakur A, Poonia B, Sarin SK, Trehanpati N, Kottilil S.Elevated hepatic lipid and interferon stimulated gene expression in HCV GT3 patients relative to non-alcoholic steatohepatitis.
2016; 10:937-946 [PMID:27193023 DOI:10.1007/s12072-016-9733-6]
22 Farquhar MJ, Humphreys IS, Rudge SA, Wilson GK, Bhattacharya B, Ciaccia M, Hu K, Zhang Q, Mailly L, Reynolds GM, Ashcroft M, Balfe P, Baumert TF, Roessler S, Wakelam MJO, McKeating JA. Autotaxin-lysophosphatidic acid receptor signalling regulates hepatitis C virus replication.
2017; 66:919-929 [PMID:28126468 DOI:10.1016/j.jhep.2017.01.009]
23 Kinast V, Leber SL, Brown RJP, Vieyres G, Behrendt P, E?bach C, Strnad P, Vondran FWR, Cornberg M, Wex C,Pietschmann T, Haybaeck J, Todt D, Steinmann E. Identification of Keratin 23 as a Hepatitis C Virus-Induced Host Factor in the Human Liver.
2019; 8 [PMID:31216713 DOI:10.3390/cells8060610]
24 Gerhard GS, Hanson A, Wilhelmsen D, Piras IS, Still CD, Chu X, Petrick AT, DiStefano JK. AEBP1 expression increases with severity of fibrosis in NASH and is regulated by glucose, palmitate, and miR-372-3p.
2019; 14:e0219764[PMID:31299062 DOI:10.1371/journal.pone.0219764]
25 Wolff G, Taranko AE, Meln I, Weinmann J, Sijmonsma T, Lerch S, Heide D, Billeter AT, Tews D, Krunic D, Fischer-Posovszky P, Müller-Stich BP, Herzig S, Grimm D, Heikenw?lder M, Kao WW, Vegiopoulos A. Diet-dependent function of the extracellular matrix proteoglycan Lumican in obesity and glucose homeostasis.
2019; 19:97-106 [PMID:30409703 DOI:10.1016/j.molmet.2018.10.007]
26 Hou W, Janech MG, Sobolesky PM, Bland AM, Samsuddin S, Alazawi W, Syn WK. Proteomic screening of plasma identifies potential noninvasive biomarkers associated with significant/advanced fibrosis in patients with nonalcoholic fatty liver disease.
2020; 40 [PMID:31860081 DOI:10.1042/BSR20190395]
27 Huang CC, Chuang JH, Chou MH, Wu CL, Chen CM, Wang CC, Chen YS, Chen CL, Tai MH. Matrilysin (MMP-7) is a major matrix metalloproteinase upregulated in biliary atresia-associated liver fibrosis.
2005; 18:941-950[PMID:15696117 DOI:10.1038/modpathol.3800374]
28 Jinawath N, Chamgramol Y, Furukawa Y, Obama K, Tsunoda T, Sripa B, Pairojkul C, Nakamura Y. Comparison of gene expression profiles between Opisthorchis viverrini and non-Opisthorchis viverrini associated human intrahepatic cholangiocarcinoma.
2006; 44:1025-1038 [PMID:17006947 DOI:10.1002/hep.21330]
29 Yovchev MI, Xue Y, Shafritz DA, Locker J, Oertel M. Repopulation of the fibrotic/cirrhotic rat liver by transplanted hepatic stem/progenitor cells and mature hepatocytes.
2014; 59:284-295 [PMID:23840008 DOI:10.1002/hep.26615]
30 Lou Y, Tian GY, Song Y, Liu YL, Chen YD, Shi JP, Yang J. Characterization of transcriptional modules related to fibrosing-NAFLD progression.
2017; 7:4748 [PMID:28684781 DOI:10.1038/s41598-017-05044-2]
31 Klag T, Thomas M, Ehmann D, Courth L, Mail?nder-Sanchez D, Weiss TS, Dayoub R, Abshagen K, Vollmar B, Thasler WE, Stange EF, Berg CP, Malek NP, Zanger UM, Wehkamp J. β-Defensin 1 Is Prominent in the Liver and Induced During Cholestasis by Bilirubin and Bile Acids
Farnesoid X Receptor and Constitutive Androstane Receptor.
2018; 9:1735 [PMID:30100908 DOI:10.3389/fimmu.2018.01735]
32 Cheung ST, Leung KL, Ip YC, Chen X, Fong DY, Ng IO, Fan ST, So S. Claudin-10 expression level is associated with recurrence of primary hepatocellular carcinoma.
2005; 11:551-556 [PMID:15701840]
33 Ding Z, Qian YB, Zhu LX, Xiong QR. Promoter methylation and mRNA expression of DKK-3 and WIF-1 in hepatocellular carcinoma.
2009; 15:2595-2601 [PMID:19496188 DOI:10.3748/wjg.15.2595]
34 Huang GW, Ding X, Chen SL, Zeng L. Expression of claudin 10 protein in hepatocellular carcinoma:impact on survival.
2011; 137:1213-1218 [PMID:21647678 DOI:10.1007/s00432-011-0987-z]
35 Bacigalupo ML, Manzi M, Rabinovich GA, Troncoso MF. Hierarchical and selective roles of galectins in hepatocarcinogenesis, liver fibrosis and inflammation of hepatocellular carcinoma.
2013; 19:8831-8849 [PMID:24379606 DOI:10.3748/wjg.v19.i47.8831]
36 Ye X, Li C, Zu X, Lin M, Liu Q, Liu J, Xu G, Chen Z, Xu Y, Liu L, Luo D, Cao Z, Shi G, Feng Z, Deng H, Liao Q, Cai C,Liao DF, Wang J, Jin J, Cao D. A Large-Scale Multicenter Study Validates Aldo-Keto Reductase Family 1 Member B10 as a Prevalent Serum Marker for Detection of Hepatocellular Carcinoma.
2019; 69:2489-2501 [PMID:30672601 DOI:10.1002/hep.30519]
37 Zhang Z, Li J, He T, Ouyang Y, Huang Y, Liu Q, Wang P, Ding J. The competitive endogenous RNA regulatory network reveals potential prognostic biomarkers for overall survival in hepatocellular carcinoma.
2019; 110:2905-2923[PMID:31335995 DOI:10.1111/cas.14138]
38 Zhang Z, Wen H, Weng J, Feng L, Liu H, Hu X, Zeng F. Silencing of EPCAM suppresses hepatic fibrosis and hepatic stellate cell proliferation in mice with alcoholic hepatitis
the PI3K/Akt/mTOR signaling pathway.
2019; 18:2239-2254 [PMID:31378124 DOI:10.1080/15384101.2019.1642067]
39 Zhang W, Ge Y, Cheng Q, Zhang Q, Fang L, Zheng J. Decorin is a pivotal effector in the extracellular matrix and tumour microenvironment.
2018; 9:5480-5491 [PMID:29435195 DOI:10.18632/oncotarget.23869]
40 Zhang G, Ezura Y, Chervoneva I, Robinson PS, Beason DP, Carine ET, Soslowsky LJ, Iozzo RV, Birk DE. Decorin regulates assembly of collagen fibrils and acquisition of biomechanical properties during tendon development.
2006; 98:1436-1449 [PMID:16518859 DOI:10.1002/jcb.20776]
41 Baghy K, Iozzo RV, Kovalszky I. Decorin-TGFβ axis in hepatic fibrosis and cirrhosis.
2012; 60:262-268 [PMID:22260996 DOI:10.1369/0022155412438104]
42 Dudás J, Kovalszky I, Gallai M, Nagy JO, Schaff Z, Knittel T, Mehde M, Neubauer K, Szalay F, Ramadori G. Expression of decorin, transforming growth factor-beta 1, tissue inhibitor metalloproteinase 1 and 2, and type IV collagenases in chronic hepatitis.
2001; 115:725-735 [PMID:11345837 DOI:10.1309/J8CD-E9C8-X4NG-GTVG]
43 Shang YK, Li F, Zhang Y, Liu ZK, Wang ZL, Bian H, Chen ZN. Systems analysis of key genes and pathways in the progression of hepatocellular carcinoma.
2018; 97:e10892 [PMID:29879025 DOI:10.1097/MD.0000000000010892]
44 Fu Y, Feng MX, Yu J, Ma MZ, Liu XJ, Li J, Yang XM, Wang YH, Zhang YL, Ao JP, Xue F, Qin W, Gu J, Xia Q, Zhang ZG. DNA methylation-mediated silencing of matricellular protein dermatopontin promotes hepatocellular carcinoma metastasis by α3β1 integrin-Rho GTPase signaling.
2014; 5:6701-6715 [PMID:25149533 DOI:10.18632/oncotarget.2239]
45 Lefebvre P, Lalloyer F, Baugé E, Pawlak M, Gheeraert C, Dehondt H, Vanhoutte J, Woitrain E, Hennuyer N, Mazuy C,Bobowski-Gérard M, Zummo FP, Derudas B, Driessen A, Hubens G, Vonghia L, Kwanten WJ, Michielsen P,Vanwolleghem T, Eeckhoute J, Verrijken A, Van Gaal L, Francque S, Staels B. Interspecies NASH disease activity wholegenome profiling identifies a fibrogenic role of PPARα-regulated dermatopontin.
2017; 2 [PMID:28679947 DOI:10.1172/jci.insight.92264]
46 Okamoto O, Fujiwara S, Abe M, Sato Y. Dermatopontin interacts with transforming growth factor beta and enhances its biological activity.
1999; 337 (Pt 3):537-541 [PMID:9895299]
47 Suda H, Yoshii D, Yamamura K, Yokouchi Y, Inomata Y. New insight into reactive ductular cells of biliary atresia provided by pathological assessment of SOX9.
2014; 30:481-492 [PMID:24658905 DOI:10.1007/s00383-014-3497-7]
48 Furuyama K, Kawaguchi Y, Akiyama H, Horiguchi M, Kodama S, Kuhara T, Hosokawa S, Elbahrawy A, Soeda T,Koizumi M, Masui T, Kawaguchi M, Takaori K, Doi R, Nishi E, Kakinoki R, Deng JM, Behringer RR, Nakamura T,Uemoto S. Continuous cell supply from a Sox9-expressing progenitor zone in adult liver, exocrine pancreas and intestine.
2011; 43:34-41 [PMID:21113154 DOI:10.1038/ng.722]
49 Yang JJ, Tao H, Deng ZY, Lu C, Li J. Non-coding RNA-mediated epigenetic regulation of liver fibrosis.
2015; 64:1386-1394 [PMID:26362725 DOI:10.1016/j.metabol.2015.08.004]
 World Journal of Gastrointestinal Oncology2022年7期
World Journal of Gastrointestinal Oncology2022年7期
- World Journal of Gastrointestinal Oncology的其它文章
- Da Vinci robot-assisted pancreato-duodenectomy in a patient with situs inversus totalis:A case report and review of literature
- Correction to “Novel long non-coding RNA LINC02532 promotes gastric cancer cell proliferation, migration, and invasion in vitro”
- Primary signet-ring cell carcinoma of the extrahepatic bile duct:A case report
- Predictors for malignant potential and deep submucosal invasion in colorectal laterally spreading tumors
- Pediatric case of colonic perivascular epithelioid cell tumor complicated with intussusception and anal incarceration:A case report
- Effect of obesity on post-operative outcomes following colorectal cancer surgery