
Three abrupt climatic events since the Late Pleistocene in the North China Plain
2013-09-27 02:37:18LinJingxingChenLeiYangMeiQuMing
Journal of Palaeogeography 2013年4期
Lin Jingxing , Chen Lei , Yang Mei Qu Ming
1.Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China
2.College of Earth Science, University of Chinese Academy of Sciences, Beijing 100049, China
3.Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences,Qingdao 266071, China
4.Jilin Second Geological Survey, Jilin 132000, China
1 lntroduction*
The data presented in this article mainly came from a study on “Quaternary marine transgressions in eastern China” (Lin and Dai, 2012).The study area includes Liaoning, Hebei, Shandong, Jiangsu, Zhejiang, Fujian,Taiwan, Guangdong, Guangxi, Hainan, Beijing, Tianjin,Shanghai, the Bohai Sea, the Yellow Sea, the East China Sea, and the South China Sea (Fig.1).The entire area approximately covers 6.25 million km2(2.65 million km2on land, 3.6 million km2in the sea).In the present study, information on three new boreholes has been added; one is from Tianjin (T1, 50 m deep), other one is from the East China Sea (E1, 56 m deep), and the third is from the South China Sea (LH19-4-1, 2630 m deep); with the addition of these boreholes, the number of boreholes has increased to 121 and the number of samples now totals 9552 (Fig.2).This research focused on the distribution pattern of foraminifera, ostracods, pollen, spores, wooly rhinoceros, elephants, water buffalo and beachrocks both in time and space.Results demonstrate that three abrupt climatic events had taken place since the Late Pleistocene in the North China Plain, which caused three great migrations of biota.
2 Three abrupt climatic events
Based on a detailed analysis of the distribution pattern of typical warm-water foraminifera in time and space, the migration of warm mammalian fauna, as well as the dispersal of cold-water ostracoda, cold mammalian fauna,and cold flora, three very warm periods and two very cold periods, termed the three abrupt climatic events (ACE),have been distinguished since the Late Pleistocene in the North China Plain.
2.1 The great migrations of the cold mammalian fauna and cold flora
Abrupt climatic changes in the Late Pleistocene causedoscillating migrations of species that could adapt to either low or high temperatures.
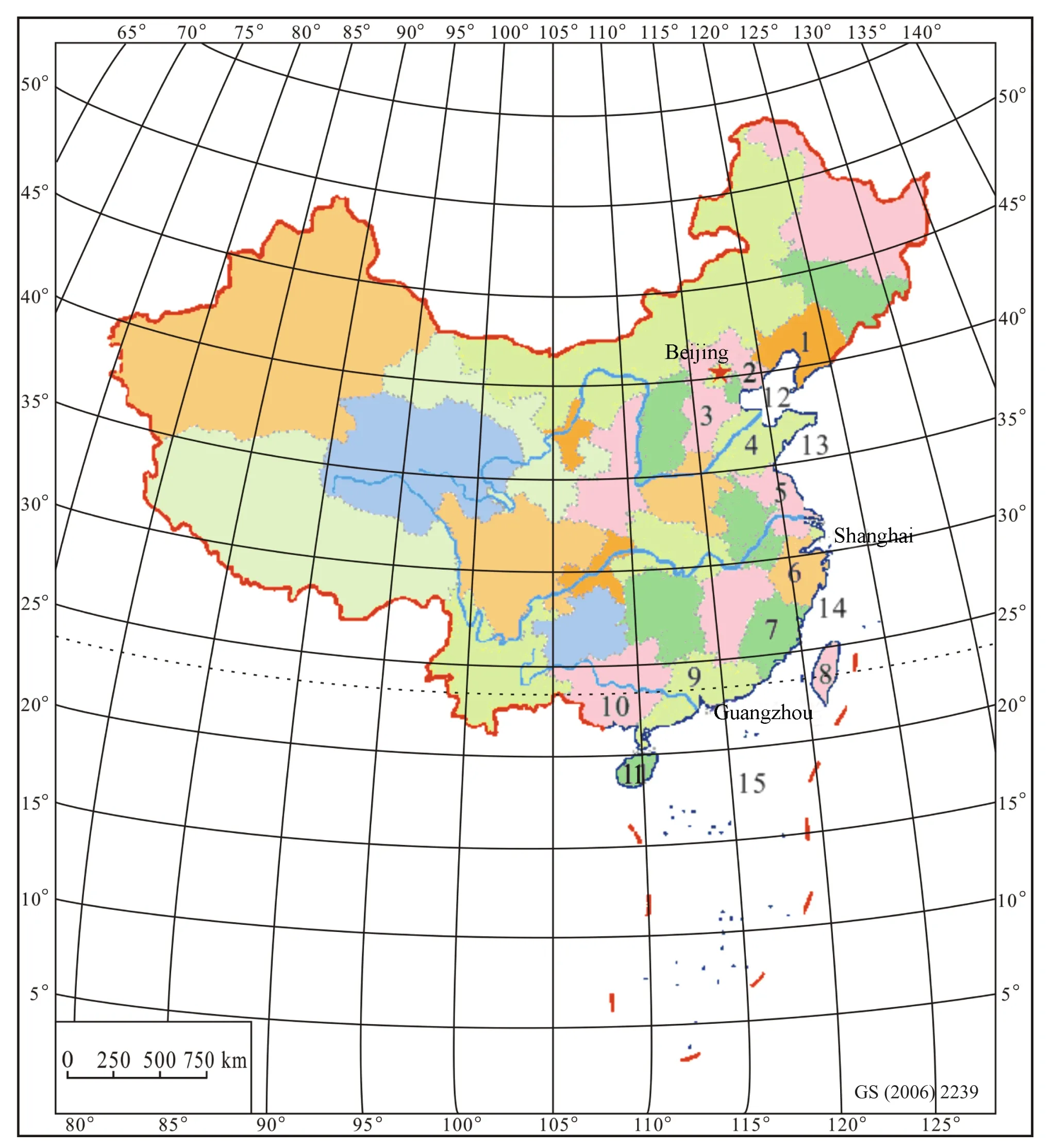
Fig.1 Study area (modified from Lin and Dai, 2012).1-Liaoning; 2-Tianjin; 3-Hebei; 4-Shandong; 5-Jiangsu; 6-Zhejiang;7-Fujian; 8-Taiwan; 9-Guangdong; 10-Guangxi; 11-Hainan; 12-Bohai Sea; 13-Yellow Sea; 14-East China Sea; 15-South China Sea.
During the two very cold periods, coldCoelodonta an?tiquitatismammalian fauna extended from North China to Qinling Mountain and the Huaihe River region in the south (Fig.3; Zhou, 1978; You and Xu, 1981).TheAbiesandPiceaflora moved from North China to the Hangzhou area and the southern Yangtze River delta region(Xuet al., 1980; Kong and Du, 1991).AbiesandPicea,which are still living today in the area of northern Beijing, moved about 1200 km, or 10° latitude from north to south (Fig.4).Considering the change of the mean temperature along a latitudinal gradient (0.7℃ per 1° latitude in eastern China), the mean temperature dropped about7℃ (at least 6℃)during that time in the North China Plain; the difference of the present annual mean temperature is about 6.4℃ between Hangzhou (18℃)and Beijing(11.6℃).
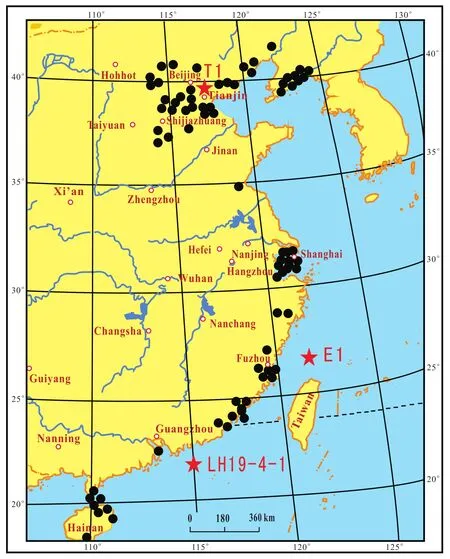
Fig.2 Geographic positions of the key ()and the three new ()boreholes in eastern China.
2.2 The great migrations of the warm mammalian fauna
During the two very warm periods in the Late Pleistocene,Elephassp.was found at Shandingdong (Fig.5),andStruthilitussp.(ostrich)was found at Malantai, both in western Beijing (Ji, 1987).The elephantPalaeoloxo?don naumaniiwas found at 13 sites in the northern Jiangsu Province (Fig.5; Fang, 1988).The water buffaloBubal?us wansijockioccurred at Salawusu, Inner Mongolia (Ji,1987).At present, elephants mainly appear in the southern part of Yunnan Province, ostriches, which disappearedfrom China occur in tropical Africa, and the water buffalo is restricted to a rough life along the north-south boundary formed by the Qinling Mountains-Huaihe River (Fig.6).During the last very warm period in the Holocene,Elephassp.occurred in the lower reaches of the Yellow River at 2490 a B.P.(Fig.5; Zhu, 1973).Three beachrock layers were found in the coastal plain of the Lushan County,Shandong Province, between 5350 a B.P.and 3600 a B.P.(Fig.5; Lin, 1985; Bi and Yuan, 1991).Beachrock can only be found in the southern part of the Xisha Islands, South China Sea, at present.All this evidence demonstrates that the warm fauna migrated in a great distance from SouthChina to North China during the warmest period since the Late Pleistocene.
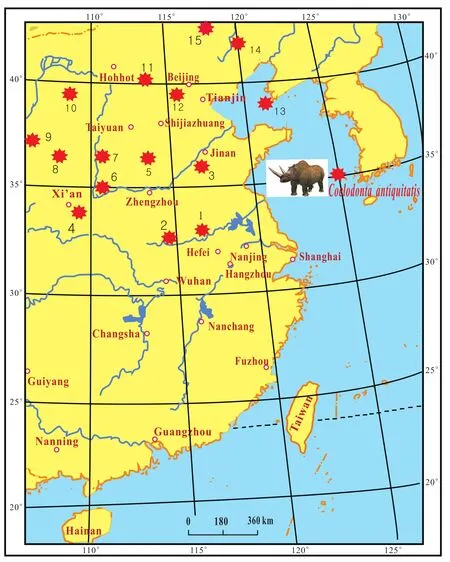
Fig.3 Distribution of Coelodonta antiquitatis during the Late Pleistocene in eastern China (data from Chow, 1978; You and Xu,1981).1-Suxian; 2-Xincai; 3-Jinan; 4-Xi’an; 5-Anyang; 6-Linyi; 7-Dingchun; 8-Salswusu; 9-Xijie; 10-Shuidonggou; 11-Nihewan; 12-Beijing; 13-Dalian; 14-Chaoyang; 15-Chifeng.
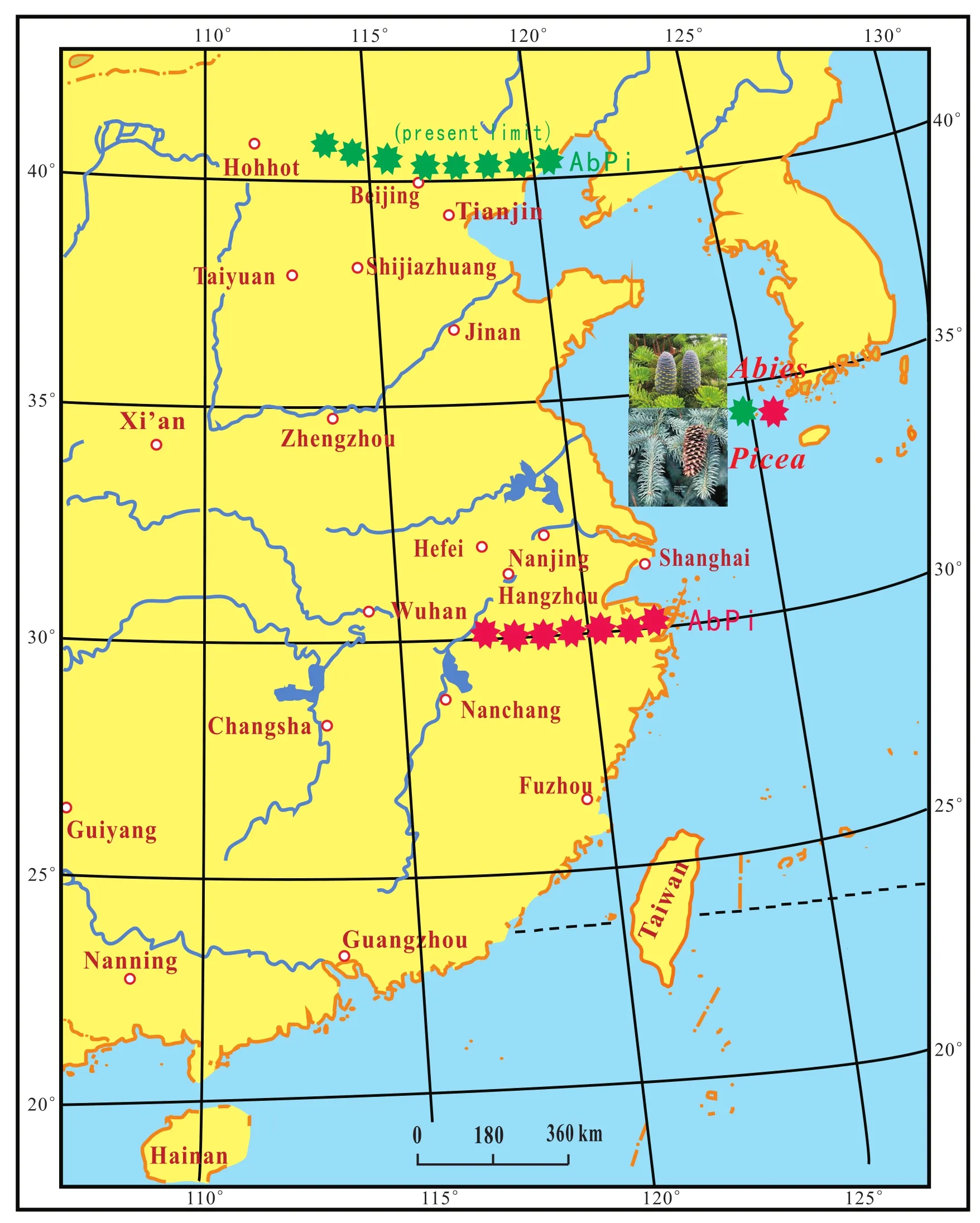
Fig.4 Southern limit of the distribution of Abies and Picea during the Late Pleistocene in eastern China (data from Xu et al., 1980;Kong and Du, 1991).-Northern boundary of Abies and Picea in the Late Pleistocene.-Present limit of Abies and Picea.
2.3 The great migrations of the typical warmwater foraminifera species group (APC)
As the migrations of the typical warm-water species group of foraminifera have been studied in great detail,we concentrate our discussion mainly on their patterns and the palaeotemperature inferences of the three warm periods which may be drawn from them (Lin, 1977; Linet al.,1989; Lin and Dai, 2012).
The typical warm-water foraminiferal species group APC at present is also called the APC species group during the ice age.Therefore, A stands for the four speciesAsterorotalia pulchella,A.venusta,A.binhaiensisandA.subtrispinos., P stands forPseudorotaliaspp.andP.schro?eteriana, and C forCellanthusspp.andC.craticulatus.

Fig.5 Distribution of the elephants Elephas sp.and Palaeoloxodon naumanii during the Late Pleistocene and Holocene in eastern China (data from Zhu, 1973; Ji, 1987; Fang, 1988).-Occurrence of Palaeoloxodon naumanii and Elephas sp.during the Late Pleistocene.-Occurrence of Elephas sp.at 2450 a B.P.during the Late Holocene.BRBRBR-Northern limit of beachrock at 5.35-3.60 a B.P.
The APC species group of foraminifera is presently living in the South China Sea and the southern part of the East China Sea.The northern limit of the species Group APC is at 26° N latitude near Fuzhou, Fujian Province (Figs.7-10).Its geographical location is the same as the northern limit of the subtropical snakeBungarus fasciatus.
The northern limit of the species Group APC during the ice age remains unknown, but considering that the mean temperature was some 6℃ lower than at present, the north-ern boundary of the species Group APC during the ice age should have shifted southward to the Palaeo-Taiwan Straits.
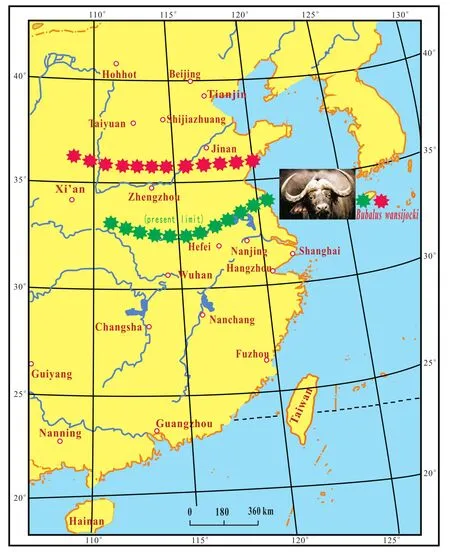
Fig.6 Northern boundary of water bufffalo Bubalus wansijocki during the Late Pleistocene in eastern China (data from Ji, 1987).-Northern boundary of Bubalus wansijocki during the Late Pleistocene.-Present-day boundary of Bubalus wansijocki.
The species Group APC dominated in the South China Sea during the Pliocene, but their northern limit during this period is still in question.There is no doubt that the species group was indigenous tropical-subtropical at this time, because they are never found outside the tropical-subtropical zone during the Pliocene.For this reason, the latitudinal shifts of the northern limit of the species Group APC can be reliably used to reconstruct the prevailing palaeotemperature of the sea.
The A and P elements of the species group spread from the South China Sea to the southern Tianjin during MTS3(MTS: Marine Transgression Stratum)in the early LatePleistocene (Fig.8; Lin and Dai, 2012).
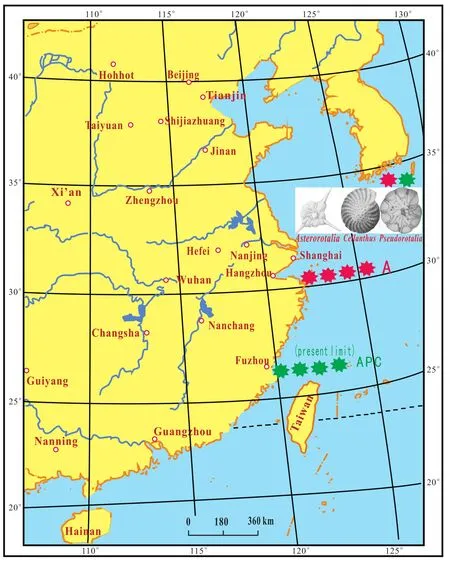
Fig.7 Northern limit of the distribution of the Asterorotalia species group (A)in the fourth marine transgression stratum (MTS4)during the late Middle Pleistocene.P-Pseudorotalia species group; C-Cellanthus species group.-Boundary of distribution of species Group A during the late Middle Pleistocene.-Present-day limit of species Group APC.
The P components migrated from the South China Sea into the Tianjin area and the components A and C extended from the South China Sea to the Shanghai area during the MTS2 in the late Late Pleistocene (Fig.9).
The C components moved from the South China Sea to the Fuding area of northern Fuzhou during the Holocene(MTS1).However, the northern limit of the components A and P remained the same during the Holocene (MTS1)as it remains today (Fig.10).
Thus, there were two times of large latitudinal shifts in the northern boundary of the species Group APC: The firstoccurred in the early Late Pleistocene, during MTS3, and probably began about 100,000 a B.P.; the second in the late Late Pleistocene, during MTS2, starting around 40,000 a B.P.During the climate optima, the species Group APC migrated from the Palaeo-South China Sea to the Palaeo-Bohai Sea; and during cold periods they moved back from the Palaeo-Bohai Sea to the Palaeo-South China Sea.
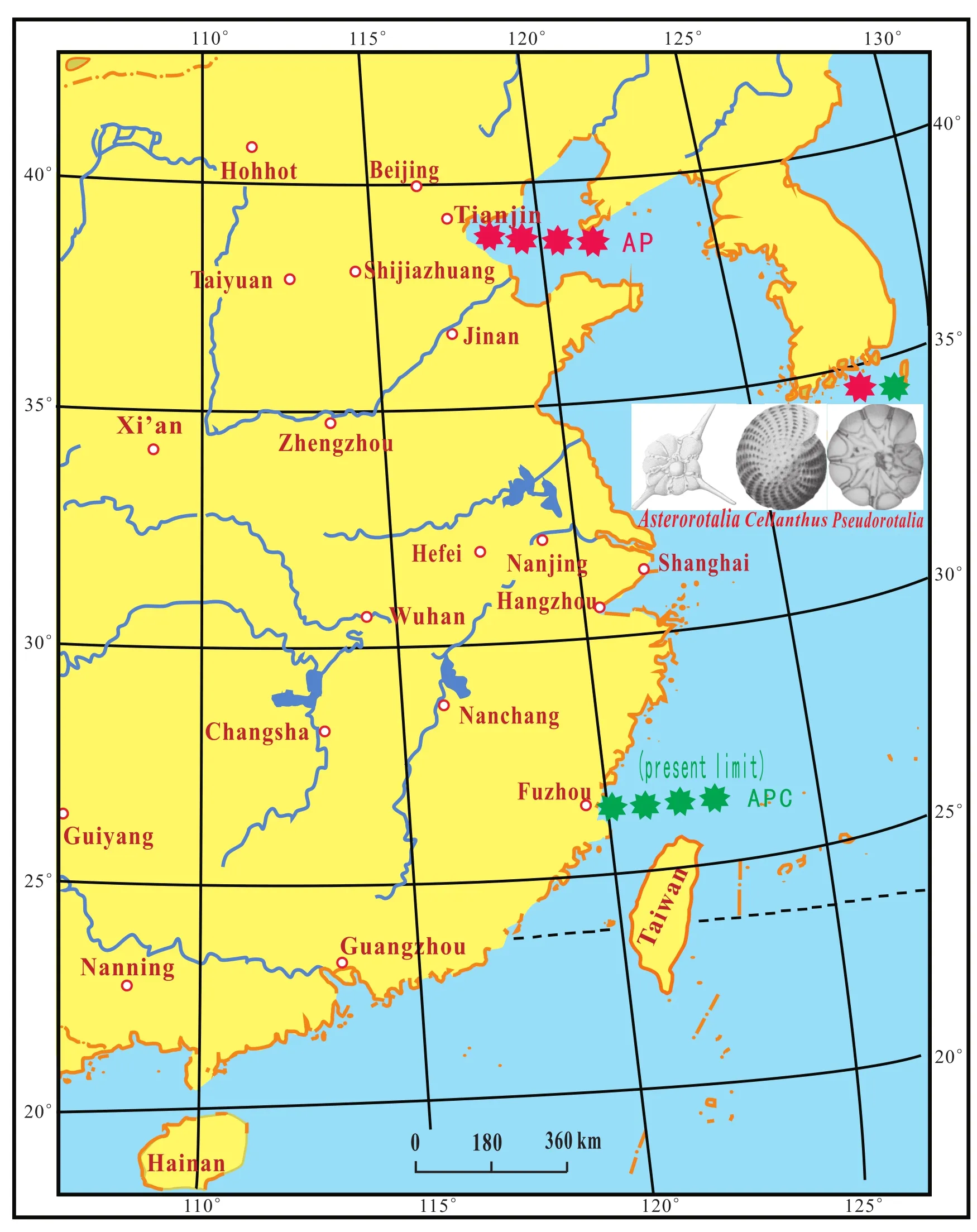
Fig.8 Northern limit of the distribution of the Asterorotalia species group (A)and Pseudorotalia species group (P)in the third marine transgression stratum (MTS3)during the early Late Pleistocene.-Distribution boundary of species Group AP during the early Late Pleistocene.-Present-day limit of species Group APC.
3 Discussion: Palaeotemperature estimation
Each species group of foraminifera is living under spe-cific environmental conditions defined by temperature,water depth and salinity, as the major factors influencing distribution.The temperature is one of the most important factors.Typical warm water organisms live only in a limited temperature range;Globorotalia tumida, for example, lives only at temperatures ranging from 18℃ to 30℃.Therefore it provides a basis for interpreting the past oceanic temperatures.
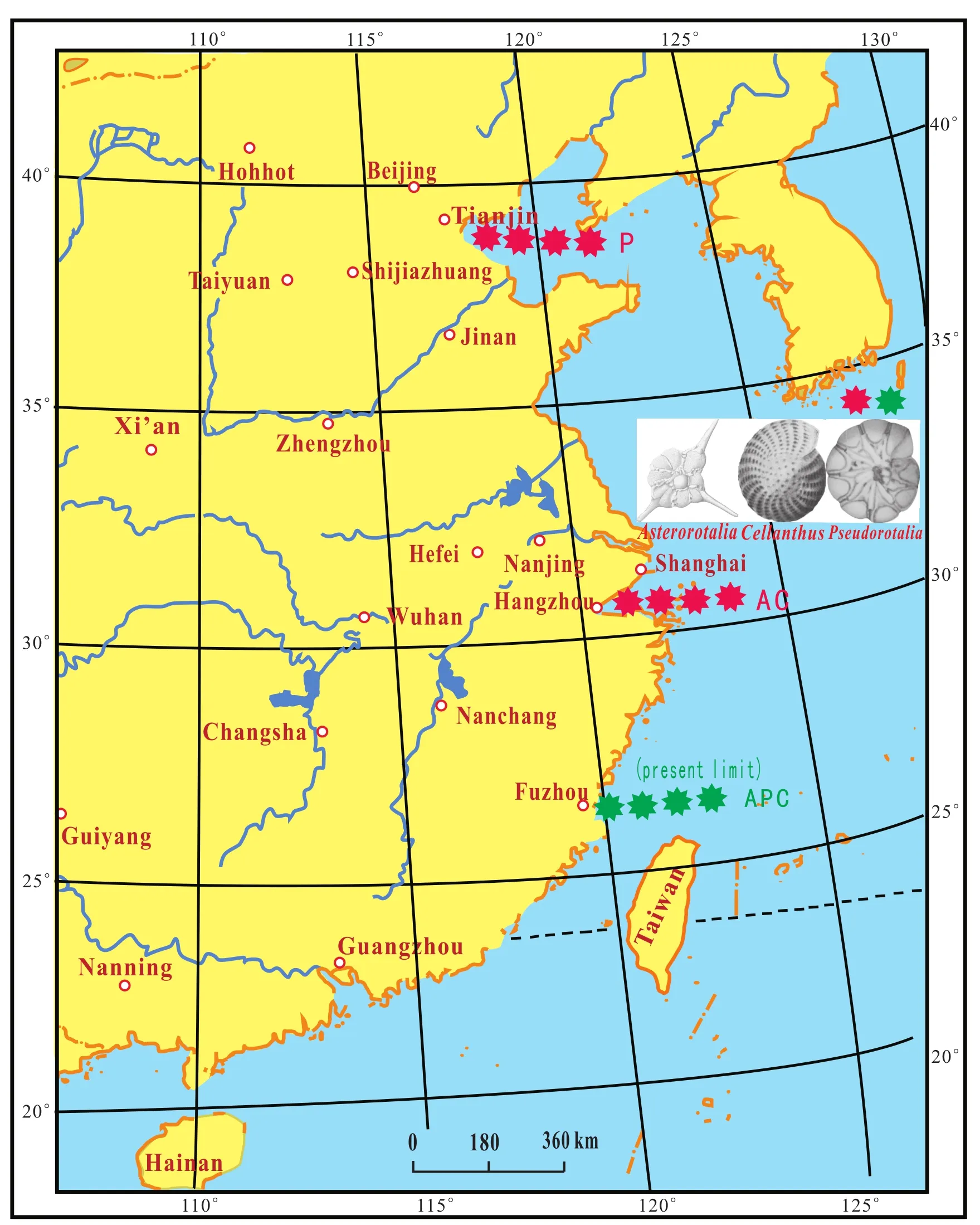
Fig.9 Northern limit of the distribution of the Asterorotalia species group (A), Cellanthus species group (C)and Pseudorotalia species group (P)in the second marine transgression stratum (MTS2)during the late Late Pleistocene.-Distribution boundary of species Group AC and species Group P during the late Late Pleistocene.-Present-day limit of species Group APC.
Studying shifts of biogeographic boundaries is one of the most important methods to estimate the past oceanic temperatures.It involves identification of a great number of foraminifera.The study of their present ecology andecological history, and the investigation of the temporal and spatial distribution of those species, can be used as temperature indicators for each geological period.Based on the study of the expansion and retreat of biogeographic distributions in the past, compared with present-day distributions, the palaeotemperature can be estimated.
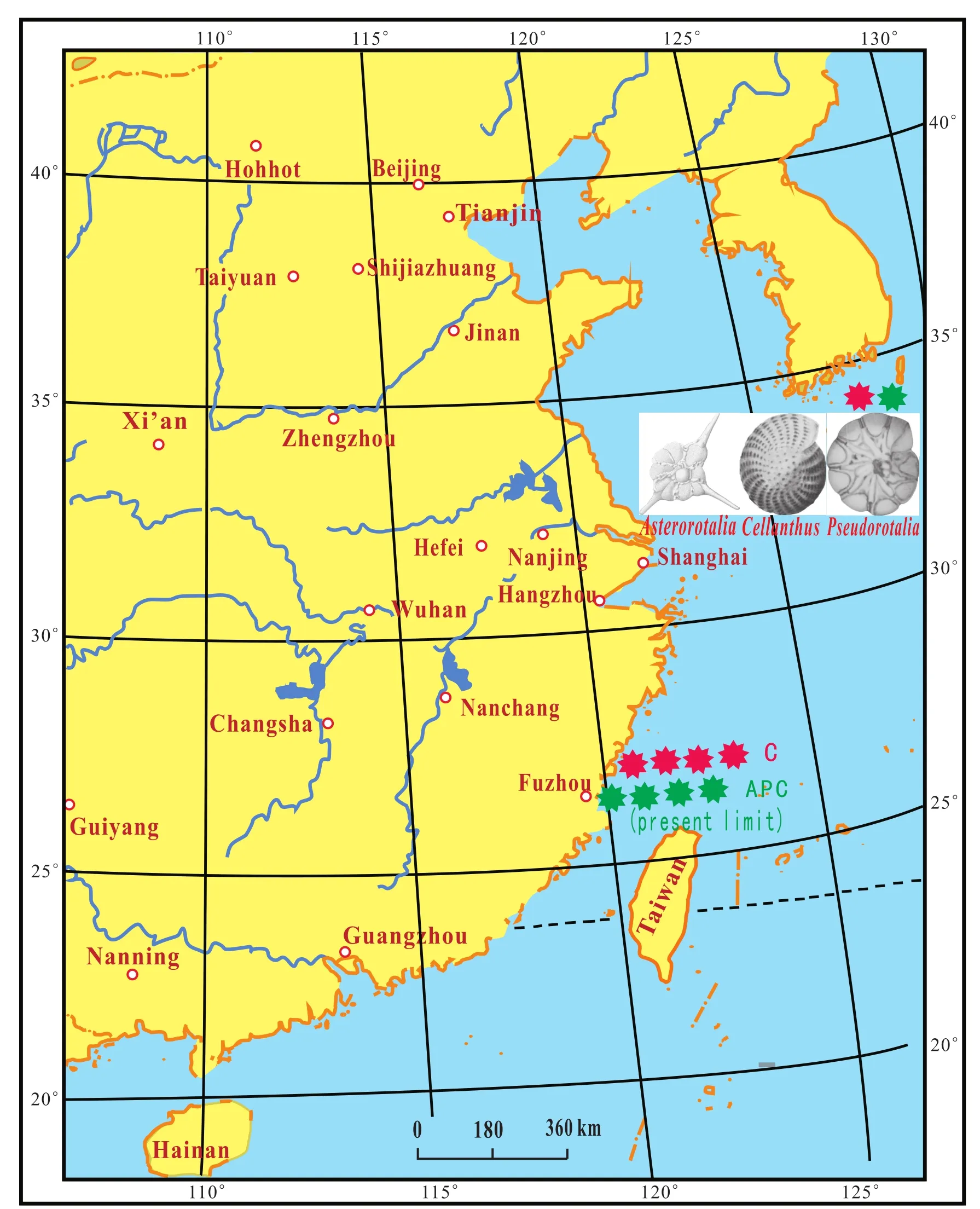
Fig.10 Northern limit of the distribution of the Cellanthus species group (C)in the first marine transgression stratum (MTS1)during the Holocene.-Distribution boundary of species Group C during the Holocene.-Present-day limit of species Group APC.
As mentioned above, the present northern boundary of the species Group APC is situated at about 26° N (near Fuzhou, Fujian Province)but during the two warmest periods of the Late Pleistocene, MTS3 and MTS2, it shifted to the Palaeo-Bohai Sea region (around 38° N).Their northern boundary thus shifted about 1400 km, correspondingto about 12° in latitude (Figs.8-10).By calculating the change rate of the mean seawater temperature per latitude(0.7℃ in eastern China), the mean temperature in the area was about 8.4℃ higher than at present.The present difference of the mean seawater temperature between Fuzhou(19.6℃)and southern Tianjin (~13℃)is about 6.6℃.Thus, the mean sea water temperature was about 7℃ higher than at present during the two warmest periods of the Late Pleistocene in the southern part of the Palaeo-Bohai Sea (Fig.11).
During the warmest period of the Middle Holocene, the palaeotemperature is estimated to be 3℃ higher than at present (judged from the northern limit of the beachrocks and the elephant fauna; Fig.11).
4 Conclusions
1)Three abrupt climatic change events happened in the North China Plain since the Late Pleistocene.
2)These abrupt climate changes caused three very warm periods and two severely cold periods.During the two warmest periods, the mean temperature was 7℃ higher than at present; and during the two extreme cold periods, the mean temperature was some 6℃ lower than today.
3)The last warm period was in the Middle Holocene,its mean temperature was 3℃ higher than at present in the North China Plain.
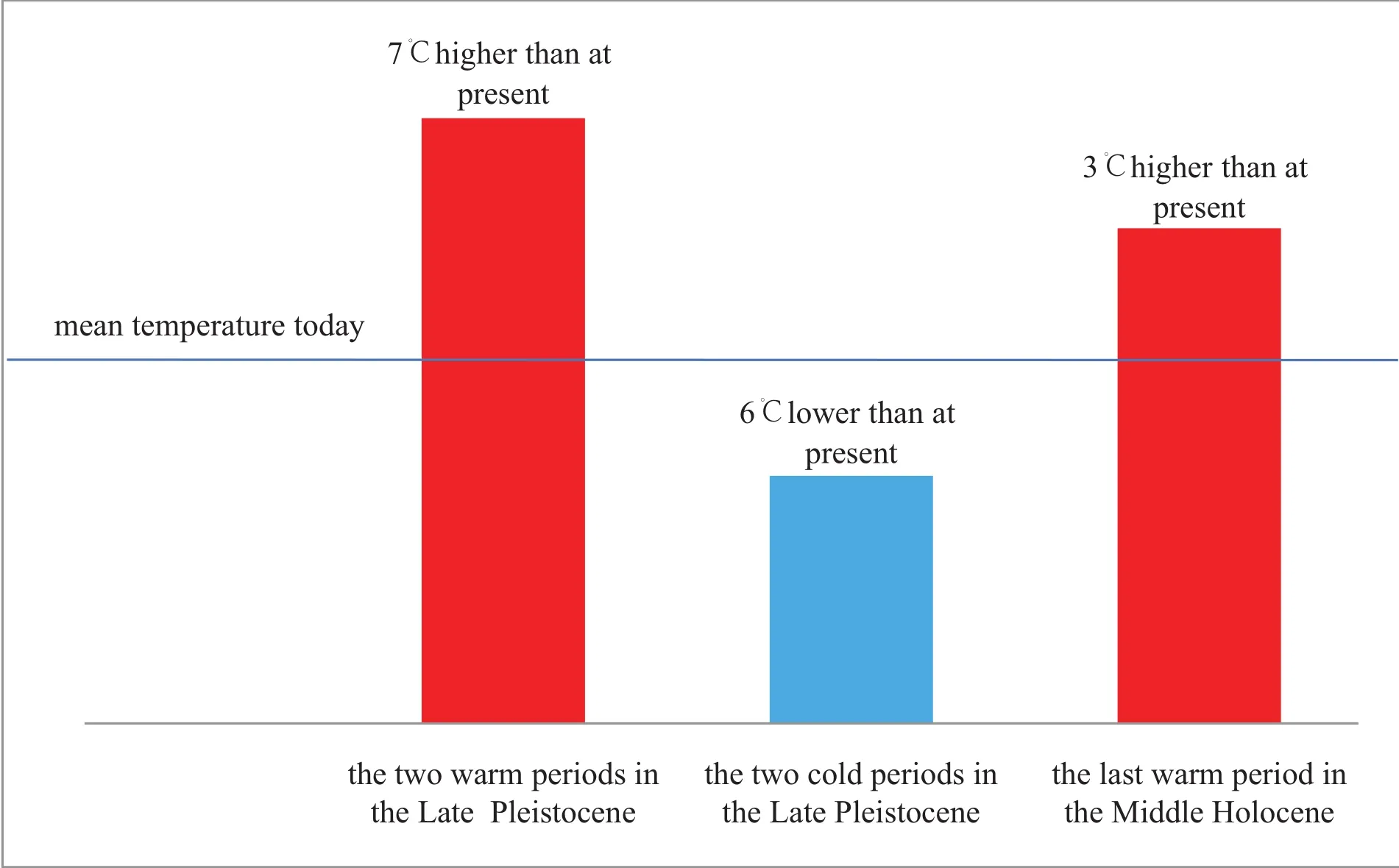
Fig.11 Mean temperature changes in North China Plain since the Late Pleistocene.
Acknowledgements
We are extremely grateful to Prof.Dr.Hao Yichun,Prof.Dr.Liu Tungsheng, Prof.Dr.Paepe R., Prof.Dr.Boski T.and Prof.Dr.Ferguson D.for their guidance,advices, comments, and critical remarks in reviewing the manuscript, which greatly improved the scientific level of the paper.
Bi Fuzhi, Yuan Youshen, 1991.Marine beachrock of Rushan, Shandong and its important significance in science.Geoscience, 2:85-91 (in Chinese with English abstract).
Fang Shangming, 1988.Elephant fossils and their horizons in the northern region of Pixian Country, Jiangsu Province.Marine Geology & Quaternary Geology, 8(1): 87-93 (in Chinese with English abstract).
Ji Hongxiang, 1987.The geographical distribution and dividing of Quaternary mammalian fauna in China.Journal of Stratigraphy,11(2): 91-102 (in Chinese with English abstract).
Kong Zhaochen, Du Naiqiu, 1991.Dramatic changes in vegetation and climate since Late Pleistocene in Eastern China.Correlationof onshore and offshore Quaternary in China.Beijing: Science Press, 165-172 (in Chinese).
Lin Baorong, 1985.The beachrock layer was found in N.China in first time.Marine Geology & Quaternary Geology, 5(4): 58 (in Chinese with English abstract).
Lin Jingxing, 1977.Preliminary notes on Quaternary transgressions and regressions in N.China Plain.Acta Geologica Sinica, 51(2):109-116 (in Chinese with English abstract).
Lin Jingxing, Dai Luping, 2012.Quaternary marine transgressions in eastern China.Journal of Palaeogeography, 1(2): 105-125.
Lin Jingxing, Zhang Shanlin, Qiu Jinbo, Wu Biaoyun, Huang Huanzhong, Huang Huizhen, Xi Jianguo, Tang Baogen, Cai Zuren, He Yubao, 1989.Quaternary marine transgression and palaeoclimate in the Yangtze River delta region.Quaternary Research, 32: 296-306 (in Chinese with English abstract).
Xu Ren, Kong Zhaochen, Du Naiqiu, 1980.Late Pleistocene Abies and Piceae flora in China and its significance in Quaternary research.Quaternary Research, 5(1): 48-56 (in Chinese with English abstract).
You Yuzhu, Xu Qinqi, 1981.The Late Pleistocene mammalian faunas of Northern China and correlation with deep-sea sediments.Vertebrata Palasiatica, 19(1): 77-86 (in Chinese with English abstract).
Zhou Benxiong, 1978.The distribution of the woolly rhinoceros and wooly mammoth.Vertebrate Palasiatica, 16(1): 45-59 (in Chinese with English abstract).
Zhu Kezhen, 1973.A preliminary study on the climate fluctuations during the last 5000 years in China.Scientia Sinica, 16(2): 226-256 (in Chinese with English abstract).
 Journal of Palaeogeography2013年4期
Journal of Palaeogeography2013年4期
- Journal of Palaeogeography的其它文章
- Deformed stromatolites in marbles of the Mesoproterozoic Wumishan Formation as evidence for synsedimentary seismic activity
- Late Quaternary palaeoenvironmental changes documented by microfaunas and shell stable isotopes in the southern Pearl River Delta plain,South China
- Taphonomy of Early Triassic fish fossils of the Vega-Phroso Siltstone Member of the Sulphur Mountain Formation near Wapiti Lake, British Columbia, Canada
- Geochemistry of the Late Paleozoic cherts in the Youjiang Basin: Implications for the basin evolution
- Milankovitch-driven cycles in the Precambrian of China: The Wumishan Formation
- Pliocene taxodiaceous fossil wood from southwestern Ukraine and its palaeoenvironmental implications