
Stress-relevant social behaviors of middle-class male cynomolgus monkeys (Macaca fascicularis)
2015-04-10 00:58:35DingCUIYuanZHOU
Zoological Research 2015年6期
Ding CUI, Yuan ZHOU
1 Key Laboratory of Behavioral Science, Institute of Psychology, Chinese Academy of Sciences, Beijing 100101, China
2University of Chinese Academy of Sciences, Beijing 100101, China
INTRODUCTION
People with different socioeconomic status are thought to experience different levels of psychosocial stress, which can lead to various health problems such as physiological and metabolic alternations, disabilities, stress-related diseases and even mortality (Adler et al, 1993, 1994; Brunner, 1997; Manuck et al, 1995; Sapolsky, 2005; Shively et al, 2005). In addition to human studies, Abbott (2003) and colleagues conducted a meta-review on dozens of nonhuman primate species, which showed strong correlations between social rank and stress level, and although the associations varied among species,certain ranks exhibited a corresponding amount of stress.Sapolsky (2005) also reviewed and reported on the close relationship between social status and health conditions in nonhuman primates, and concluded that social status markedly influenced health. Accordingly, investigations on social status and stress levels have drawn increasing attention.
Bupa claims that more than a half of middle-class managers suffer from overwhelming stress problems and experience mental health conditions such as depression (Bupa Research,2013). Similarly, Edwards et al (2013) found that middle-class barbary macaques experience more social stress than those with relatively higher or lower social status. Thereby, middleclass stress seems to draw increasing attentions from general studies of graded variations between social rank and stress conditions. However, the origins of middle-class stress and how middle-ranked individuals adapt to their particular social status remain poorly understood.1
In the present study, we selected beta monkeys in captive,male-only, cynomolgus monkey groups (as middle-ranked individuals), and observed and analyzed four types of social behaviors (aggression, avoidance, grooming and mounting) in the hope of clarifying the origins of middle-class stress and how beta monkeys adapt to their social status from a behavioral point of view.
MATERIAL AND METHODS
Subjects
Data were collected from seven isosexual social colonies of male cynomolgus monkeys (Macaca fascicularis), with 10-15 individuals aged 4-5 years in each group. The monkeys were housed in colonies (3 m×5 m×3 m) at Hainan Jingang Biotech Co., Ltd. (Tian & Ma, 2014), and had free access to water and monkey chow supplemented with fruit and vegetables. The animals were reproductively intact. The members in each group were unchanged for at least two years prior to initial sampling.The treatment of the animals was conducted in strict accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and was approved by the Biological Research Ethics Committee of the Hainan Jingang Biotech Co., Ltd.
Behavioral sampling
The monkeys were familiarized with observers until they were completely adapted to observers remaining in front of the cage.Observers stood or sat at a distance of at least 2.5 m from the cage. Except for taking sampling notes, observers did not move about freely or feed the animals. Actual behavior sampling started after the adaptation period.
Social status was determined via instant scan sampling(Altmann, 1974) during the adaptation period, and was based on the order of eating and the ability to defeat others in each group. More specifically and theoretically, the animal that defeated all other members in its group and ate first when food was provided was ranked as the alpha (top) male; the animal that defeated all but the alpha male was ranked as the beta(second) male; and so forth. In the present study, the top two ranked monkeys in each group were very clear, and their respective statuses were stable during the observation period.However, the ranks of the remaining individuals (excluding the lowest) were unclear and unstable because their observable behavioral differences were difficult to determine. Similar to human middle-ranked managers (Bupa Research, 2013), beta males had to obey the alpha males (boss) as well as maintain their beta status over the other group members. We therefore selected the middle-ranked beta males as the target individuals in this study as they were hierarchically positioned between the alpha and lower-ranked members.
The frequencies of social behaviors, including aggression,avoidance, grooming and mounting, were recorded using focal animal sampling (Altmann, 1974). The frequency of social behaviors between the beta male and other cage mates was recorded. From the action direction point of view, we recorded six sub-categories for each social behavioral type, namely,actions initiated by the alpha male toward the beta male, beta male toward the alpha male, alpha male toward ‘others’, ‘others’toward the alpha male, beta male toward ‘others’, and ‘others’toward the beta male. Each monkey group was recorded for three consecutive days in total, six times a day, 10 min per time between 1700h and 1900h.
Statistical analysis
Student’s t-tests (paired) were used for comparison of the frequency (over entire observation period) differences among sub-categories in each social behavioral category. The t-tests were two-tailed withP<0.05 considered to be statistically significant.
RESULTS
Aggressive behavior
No aggressive behavior, across all seven monkey groups and the entire observation period, was initiated by the beta male or‘others’ toward the alpha male, nor from ‘others’ toward the beta male. Aggression mainly occurred between the alpha male and beta male/‘others’, as well as the beta male and ‘others’. As shown in Table 1, the average frequency of the alpha male attacking the beta male or ‘others’ was 3.14 and 2.57,respectively. The beta males received more than half (55%, 22 out of 40 occurrences) the aggressive actions initiated by the alpha males.
In addition, compared with the alpha male, the beta male attacked the ‘others’ more frequently (average frequency of 2.57 v.s. 7.00, P=0.0008, t=6.16, SD=1.9). The beta male was attacked by the alpha male on 72.7% of occasions (16 out of 22 times per group), after which the beta male immediately initiated aggressive behavior toward the ‘others’. By comparing the aggressive behaviors received (A to B, 3.14 times per group on average) and given by the beta male (B to ‘others’, 7.00 times per group on average), the beta male was more often an aggressor than victim (P=0.006,t=4.12,SD=2.47).

Table 1 Frequencies of aggressive behavior
Avoidance behavior
Avoidance behavior occurred more frequently than the other behaviors, as seen from the total number of each sub-category(97, 68, and 89 times) shown in Table 2. The beta male avoided the alpha male more frequently than the ‘others’ did (13.86v.s.9.71 times per group on average,P=0.018,t=3.19,SD=3.43).There was no significant avoidance predilection of ‘others’ to either the alpha or beta males, although they showed more, intotal number (68v.s.89 times), avoidance behavior toward the beta males. The alpha males exhibited no avoidance behavior.

Table 2 Frequencies of avoidance behavior
Grooming (affinitive behavior)
As shown in Table 3, grooming was dual directional among the alpha male, beta male and ‘others’. The beta males groomed the alpha males much more frequently than the reverse (5.43 v.s. 1.00 times on average, P=0.003, t=4.55, SD=2.57). The beta males also groomed the alpha males much more frequently than they did the ‘others’ (5.43v.s.1.71 time on average, P=0.003, t=4.6, SD=2.14). We did not find any significant differences in the beta male receiving or providing grooming from/to the ‘others’. This differed from that observed between the alpha males and ‘others’, namely, alpha males groomed the ‘others’ much less frequently than ‘others’groomed the alpha males (0.71 v.s. 2.29 on average, P=0.017,t=3.27,SD=1.27).

Table 3 Frequencies of grooming behavior
Mounting behavior
Mounting was the least frequently performed behavior among the four categories of social behaviors. As shown in Table 4, the alpha males mounted the beta males a total of five times, with beta males only mounting an alpha male on one occasion in one group, but mounting the ‘others’ on seven occasions. No mounting behaviors were observed from the ‘others’ toward either the alpha or beta males. No significant differences were observed between any two sub-categories in regards to mounting behavior.
DISCUSSION
In the present study, we investigated two aspects relevant to the stress conditions of beta individuals in male-only cynomolgus monkey groups; namely, the origins of stress experienced by beta males and the adaptation of beta males to stress from their own social status. Observations of various social behavioral contacts between the beta male and his cage mates were conducted to clarify these questions from a behavioral point of view.
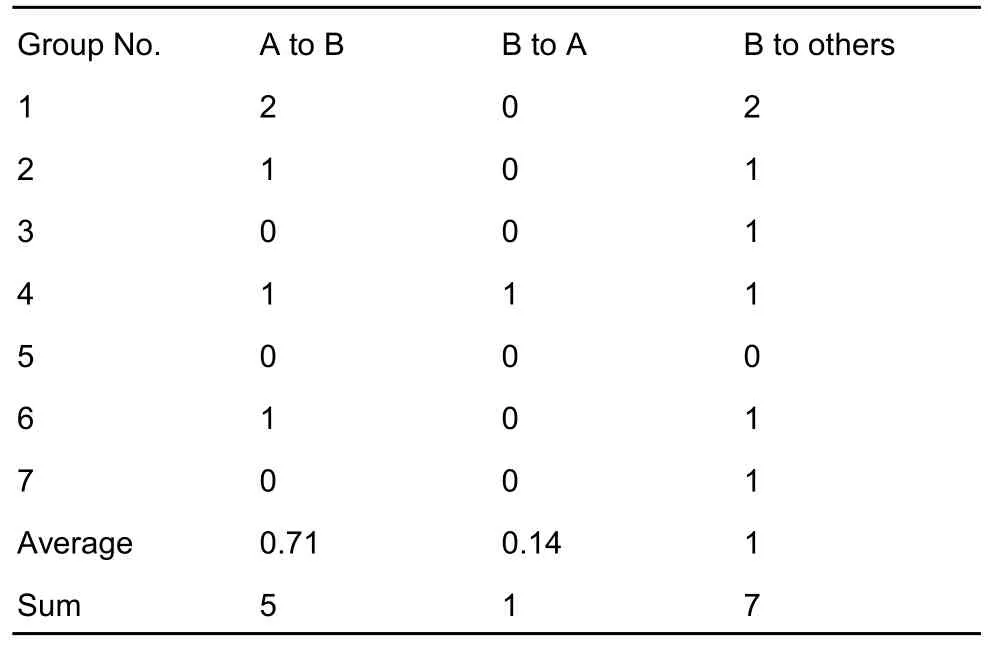
Table 4 Frequencies of mounting behavior
We suggested that the origin of stress for the beta male was mainly from being attacked by the alpha male. According to our results, the beta male received 55% of aggressive behavior initiated by the alpha male. As a single target, the beta male received more aggression from the alpha male than the total number of attacks on the other 8-13 individuals. This strongly suggests that the beta male was the main target of aggression from the alpha male. Furthermore, a different research group in our lab found a significant positive correlation between aggressive behavior and hair cortisol (a stress hormone)concentration in adult male rhesus monkeys (unpublished data);namely, the more aggression an individual received, the higher their hair cortisol concentrations, and thus the higher their stress levels. Our results indicated that beta male stress mainly originated from aggressive pressure from the alpha male in each respective group.
Individuals with a certain social status have to adapt to their specific situations, including playing their expected role and dealing with stress. As stated previously, middle-ranked human managers have to obey their higher-ranked bosses, while also leading their lower-ranked subordinates, and thereby hold special status and high levels of stress. Similarly, in the present study, the beta male held a ‘middle-class’ rank, thus while obeying the alpha male, he also had to maintain his beta status in the group. How do beta males adapt to the stress of middle class? We found 72.7% of aggressions when the beta male was attacked by the alpha male, after which the beta male immediately initiated aggressive behavior toward the ‘others’.The beta male also attacked ‘others’ more frequently than the alpha male did (7.00 v.s. 2.57 times per group on average,P=0.0008,t=6.16,SD=1.9). The intense aggressiveness of the beta male (similar rate as that of alpha males in total, 49 v.s. 40;and more frequently to ‘others’ than that of the alpha males, 49v.s.18,P=0.0008,t=6.16,SD=1.9) was likely performed to maintain his social status as the ability to defeat group members was a determinant of social rank; on the other hand,this aggressive behavior (as suggested from the 72.7%redirection of aggression by the beta male) may be a way to release high tension from being attacked by the alpha male.Previous studies have reported that social behaviors such as grooming, avoidance and mounting can serve as important functions to reduce the stress of opponents, as well as to help decrease the frequency of conflicts (i.e., Bernstein & Ehardt,1985; Castles & Whiten, 1998; Cords, 1992; Das et al, 1998;Faraut et al, 2015). In particular, affinitive and reconciliation behaviors play a significant role in soothing stress and reducing conflict. Many studies suggest that to maintain a relatively balanced and peaceful social environment, after fighting or threatening one another, most animals show friendliness by touching, hugging or grooming one another (Aureli et al 1989;Cheney & Seyfarth, 1989; De Waal & Roosmalen, 1979; De Waal & Yoshihara, 1983; York & Rowell, 1988). Compared with the ‘others’, we found that beta males showed more avoidance behavior toward the alpha males (13.86 v.s. 9.71 times per group, on average,P=0.018,t=3.19,SD=3.43), and groomed the alpha male more frequently (5.43 v.s. 1.71 times, on average, P=0.003, t=4.6, SD=2.14). We considered these beta male avoidance and grooming behaviors as strategies to relieve stress from aggression and reduce potential conflicts with the alpha male. In addition, with no significant differences in frequencies (1.71 v.s. 2.00 times per group on average), the grooming behaviors between beta and ‘others’ could also help with soothing stress of the beta male. Furthermore, although mounting behaviors between the beta and alpha males (six times in total)/’others’ (seven times in total) were rare, they may have served to mitigate stress as well.
It is important to note that certain factors that may contribute to stress, such as breeding and survival issues, were controlled in our observation groups, which might also contribute to answering the questions of interest. Specifically, all monkey subjects had free access to food and water, which eliminated the stress for survival. They also had no visual contact with females, which reduced the effects of reproduction and breeding season. Therefore, beta male stress was most likely from social interactions with their male cage mates.
In conclusion, due to its middle rank within the hierarchical social group, the beta males in the captive male-only cynomolgus monkey groups experienced a special stressful situation due to aggressions from the higher-ranked alpha males. To reduce their stress and adapt to their beta status, the beta males were actively involved in social behaviors, i.e.,aggression toward ‘others’ after being attacked, as well as receiving and preforming avoidance, grooming and mounting behavior. Further physiological studies, such as comparison of plasma/hair cortisol levels among individuals holding different social statuses, should be conducted in combination with future behavioral analysis.
Abbott DH, Keverne EB, Bercovitch FB, Shively CA, Mendoza SP, Saltzman W, Snowdon CT, Ziegler TE, Banjevic M, Garland T, Sapolsky, RM.2003. Are subordinates always stressed? A comparative analysis of rank differences in cortisol levels among primates.Hormones and Behavior,43(1): 67-82.
Adler NE, Boyce T, Chesney MA, Cohen S, Folkman S, Kahn RL, Syme SL.1994. Socioeconomic status and health: The challenge of the gradient.American Psychologist, 49(1): 15-24.
Adler NE, Boyce WT, Chesney MA, Folkman S, Syme SL. 1993. Socioeconomic inequalities in health: No easy solution.JAMA-the Journal of the American Medical Association, 269(24): 3140-3145.
Altmann J. 1974. Observational study of behavior: Sampling methods.Behaviour, 49(3): 227-67.
Aureli F, Van Schaik CP, Van Hooff JA. 1989. Functional aspects of reconciliation among captive long‐tailed macaques (Macaca fascicularis).American Journal of Primatology, 19(1): 39-51.
Bernstein IS, Ehardt CL. 1985. Agonistic aiding: Kinship, rank, age, and sex influences.American Journal of Primatology, 8(1): 37–52.
Brunner E. 1997. Socioeconomic determinants of health: stress and the biology of inequality.British Medical Journal, 314(7092): 1472-1476.
Bupa Research. 2013. Managers too stressed to notice junior staff struggle.Retrieved from http://www.bupa.com/media-centre/press-releases/uk/22-nov-2013-managers-too-stressed-to-notice-junior-staff-struggle/.
Castles D, Whiten A. 1998. Post-conflict behaviour of wild olive baboons. I.Reconciliation, redirection and consolation.Ethology, 104(2): 126-147.
Cheney DL, Seyfarth RM. 1989. Redirected aggression and reconciliation among vervet monkeys,Cercopithecus aethiops.Behaviour, 110(1): 258-275.
Cords M. 1992. Post-conflict reunions and reconciliation in long-tailed macaques.Animal Behaviour, 44(1): 57-61.
Das M, Penke Z, van Hooff JARAM. 1998. Postconflict affiliation and stress-related behavior of long-tailed macaque aggressors.International Journal of Primatology, 19(1): 53-71.
De Waal FB, Roosmalen A. 1979. Reconciliation and consolation among chimpanzees.Behaviour Ecology Sociobiology, 5(1): 55-66.
De Waal FB, Yoshihara D. 1983. Reconciliation and redirected affection in rhesus monkeys.Behaviour, 85(3): 224-241.
Edwards KL, Walker SL, Bodeham RF, Ritchie H, Shultz S. 2013. Associations between social behaviours and adrenal activity in female Barbary macaques: Consequences of study design.General and Comparative Endocrinology, 186: 72-79.
Faraut L, Northwood A, Majolo B. 2015. The functions of non-reproductive mounts among male Barbary macaques (Macaca sylvanus).American Journal of Primatology, 77(11):1149–1157.
Manuck SB, Marsland AL, Kaplan JR, Williams JK. 1995. The pathogenicity of behavior and its neuroendocrine mediation: An example from coronary artery disease.Psychosomatic Medicine, 57(3): 275-283.
Sapolsky RM. 2005. The influence of social hierarchy on primate health.Science, 308(1): 648-652.
Shively CA, Register TC, Friedman DP, Morgan TM, Thompson J, Lanier T.2005. Social stress-associated depression in adult female cynomolgus monkeys (Macaca fascicularis).Biological Psychology, 69(1): 67-84.
Tian CY, Ma YY. 2014. Involve both genetic and environmental factors to build monkey models of mental disorders.Zoology Research, 35(3): 170-171.
York AD, Rowell TE. 1988. Reconciliation following aggression in patas monkeys,Erythrocebus patas.Animal Behaviour, 36(2): 502-509.

- Zoological Research的其它文章
- Morphometric studies of genus Placocheilus (Teleostei:Cypriniformes) from Red River, China
- Patterns of reptile and amphibian species richness along elevational gradients in Mt. Kenya
- Morphometric variability of Arctodiaptomus salinus(Copepoda) in the Mediterranean-Black Sea region
- Accelerated evolution of constraint elements for hematophagic adaptation in mosquitoes
- Physiological approaches to understanding molecular actions on dorsolateral prefrontal cortical neurons underlying higher cognitive processing
- Editor’s comments