
Predatory Efficacy of Cotton Inhabiting Spiders on Bemisiatabaci, Amrascadevastans Thripstabaci and Helicoverpa armigera in Laboratory Conditions
2015-12-29 01:22:35MohsinLiYijingTangLijieMaqsoodTingMasunHanLemengKhalilAndleebMuhammadandSaleem
Mohsin S B, Li Yi-jing, Tang Li-jie, Maqsood I, Ting Ma-sun, Han Le-meng, Khalil U R, Andleeb S, Muhammad S K, and Saleem M A
1College of Veterinary Medicine, Northeast Agricultural University, Harbin 150030, China
2College of Environment Resources, Northeast Agricultural University, Harbin 150030, China
3Department of Zoology Wildlife and Fishery, Government College University Faisalabad, Pakistan
Predatory Efficacy of Cotton Inhabiting Spiders on Bemisiatabaci, Amrascadevastans Thripstabaci and Helicoverpa armigera in Laboratory Conditions
Mohsin S B1, Li Yi-jing1, Tang Li-jie1, Maqsood I1, Ting Ma-sun1, Han Le-meng1, Khalil U R2, Andleeb S2, Muhammad S K3, and Saleem M A3
1College of Veterinary Medicine, Northeast Agricultural University, Harbin 150030, China
2College of Environment Resources, Northeast Agricultural University, Harbin 150030, China
3Department of Zoology Wildlife and Fishery, Government College University Faisalabad, Pakistan
Distinction of predator's diet and prey choice preference is a hot topic of current investigations. Spider being generalist predator and cosmopoliter in nature acts as biological control agent in many agro-ecosystems. In the current study, predatory efficacy of five spiders (Pardosa birmanica, Cyclosa insulana, Thomisus projectus, Plexippus paykulli and Lycosa terrestris) inhabiting the cotton fieldevaluated in the laboratory conditions maintained at (27±2)℃ temperature, 65%±5% relative humidity and 12 : 12 h of light and dark photoperiod. Four key cotton pests' viz. Bemisiatabaci, Amrascadevastans Thripstabaci and Helicoverpa armigera were utilized in the choice and no-choice predatory studies. The findings of current investigation revealed that each predatory spider utilized at least one pest species. Predation rate was found higher in the no-choice predation because of unavailability of substitutes feeding source. The investigation also indicated each predatory spider killed more pests than consumed. The findings of this study support the predatory role of spiders in controlling the cotton major pests.
predatory, spider, efficacy, cotton pest, laboratory, condition
Introduction
Cotton being important cash crop played a vital role in the economy of Pakistan since partition. It is perhaps cultivated in all the regions of world in variety of climate. The varied climate pattern favors a variety of pests' attacks reducing the crop yield (Channa et al., 2006). Traditional methods like chemical control are being employed which may also disturb the natural ecosystem by killing the natural enemies of the pest along with pests species. Moreover, the repeated use of pesticides also develops resistance in the pests against various groups of pesticides.
In the current scenario, interest is being develop towards integrated pest management (IPM) to diminish reliance on only chemical methods (Butt and Sherawat, 2012; Khuhro et al., 2012). Spider being voracious predator and cosmopolite in nature form one of the important biological control agent (Khurro et al., 2012). It dominantly inhibits the cotton fields with 3 935 genera and 44 906 described species (Platnick 2014; Sharma, 2014). Pray searching ability, ease of multiplication and polyphagy in nature attributes the role as potential predator in the biological control (Rajeswaran et al., 2005) controllingthe aphids, mites of cotton (Khuhro et al., 2012), leafhopper (Jeyaparrathi et al., 2013), lepidopteron (Hooks et al., 2003) of various agro-ecosystems.
Unfortunately, predatory spiders are very limitedly applied in the biological studies, so the current study aimed to investigate the predatory efficacy of five predatory spiders against four key pests in the laboratory conditions.
Materials and Methods
Collection of spiders and pests
The predatory spiders were collected by hand picking method brought in the laboratory identified upto species level using the keys formulated by Tikader and Biswas (1981), Tikadar and Malhota (1980), Barrion and Litsinger (1995) and other relevant literature, sorted the sex wise and transferred to cage for predatory studies. The identified spiders were washed with xylene and preserved in vials, containing 70% ethanol few drops of glycerin and used as reference for further identification.
The pests were collected by using self-made aspirator (Fig. 1A), visual searching and hand picking method from cotton fields kept alive to use in the experiment. Adult Jassid, Thrips and white fly were used in the predatory assay. Only 1st instar of American bollworm was utilized in the study and reared in the laboratory conditions on the artificial diet formulated by Gupta et al. (2004) with some modifications provided in Table 1.
Rearing of H. armigera (Hubner)
Adult moths of both sexes were collected from the collected from fields of Ayub Agricultural Research Institute, NIAB and vicinity of Faisalabad in plastic vial. Each vial was capped with a finely perforated mesh to allow the air. The moths of equal number from both sexes were transferred to glass chimneys having muslin cloth on both openings. A Petri-dish was placed at the bottom of the chimney. Ten to 15 pairs of moths were presented in single chimney, and provided with 10% sugar solution as feed present in plastic vial having cotton pad (Fig. 1B). Sugar solution soaked the cotton pads by capillary action and moths sucked the solution from cotton pads. Eggs laid by the female moths were collected daily from muslin cloth present on the openings of the chimney and were transferred in plastic bags. These bags were labeled with date and kept for incubation and maintained at (27 ± 2)℃, 65% ± 5% RH and a 12 : 12 (L : D) photoperiod.

Fig. 1 A, Self-made aspirator for collection of pests; B, Pupuational chamber used for development of pupae to adult moth; C, Rearing tray for rearing of 3rd to 6th istars of american bollworm; D, Glass chimney used for mating of adult moths.

Table 1 Ingredients of artificial diet
The healthy newly emerged larvae of 1st instar were used in the predatory studies where remaining were transferred in large sized Petri-dish in which pieces of artificial diet were presented. This was placed in a box covered with black cloth. Up to 3rd instar, larvae remained in the Petri-dish and then transferred into rearing trays (30 cm×15 cm×4.5 cm) provided with artificial diet (Fig. 1C). Each rearing tray was divided into 24 squared chambers having lid with holes (diameter: 2 cm) covered with brass screen for ventilation. The larvae were placed in separate chambers (one larva in each) to avoid cannibalism. The chambers were cleaned daily to remove the feces of larvae. Pupae were allowed to form from the larvae which were transferred in pupal chamber (Fig. 1D) and the whole cycle was repeated.
Predatory studies
To determine the feeding potential of predatory spiders, four different sucking insect pests' viz. Jassid (A. devastans), white fly (B. tabaci), thrips (T. tabaci) and bollworm (H. armigera) were tested in choice and no choice feeding. Initially, 20 specimens of each live pest were put in the large Petri-dishes on fresh cotton leaves. Later, 24 h starved spiders were released using camel hair brush. Five replications were kept for determining the feeding efficiency of each predator species which were collected from the same cotton plant considering the same age of predators. In no choice feeding 20 adults of each Jassid, white fly and thrips were provided separately to a single predator for 24 h. In the similar way, 20 larvae of American bollworm were provided to the each predator. Pests fed or killed by each predatory spider were counted after 24 h.
Analysis of data
A complete randomized designed was used with five replicates in each case. Data was presented in mean ± SD and analyzed through Duncan's multiple range test using IBM SPSS v.20.
Results
In this study, five predatory spiders viz, P. birmanica, C. insulana, T. projectus, P. paykulli and L. terretris were tested against four key pests white fly, Jassid (B. tabaci) (A. devastans), thrips (T. tabaci) and American bollworm (H. armigera) in the laboratory conditions at (27 ± 2)℃, 65% ± 5% RH and a 12 : 12 (L : D) photoperiod. Feeding potential and host preference was recorded on the daily basis after 24 h. Table 2 provided the rate of feeding and host preference in the choice feeding. Each spider showed different feeding preferences, such as P. birmanica preferred to feed more on Jassid (8.40 ± 2.30) than other. C. insulana efficiently consumed white fly (11.67 ± 1.75) perhaps due to same ecological habitats the leaves of plants. T. projectus not only fed on white fly (8.40 ± 1.95), but also showed preference towards American bollworm (5.4 ± 0.90). P. paykulli was the only species that killed and consumed the live thrips (8.8 ± 1.79) more efficiently than other spider species. L. terretris consumed white fly (11.20 ± 1.64) moreactively then Jassid (5.75 ± 1.96) and finally thrips (2.2 ± 0.45) and American bollworm (2.2 ± 0.84) at the same rate. Being more active, female spiders showed slightly higher predation potential than male spiders. The preference for consuming the specific pests was found for the male spiders in feeding test (Table 3).
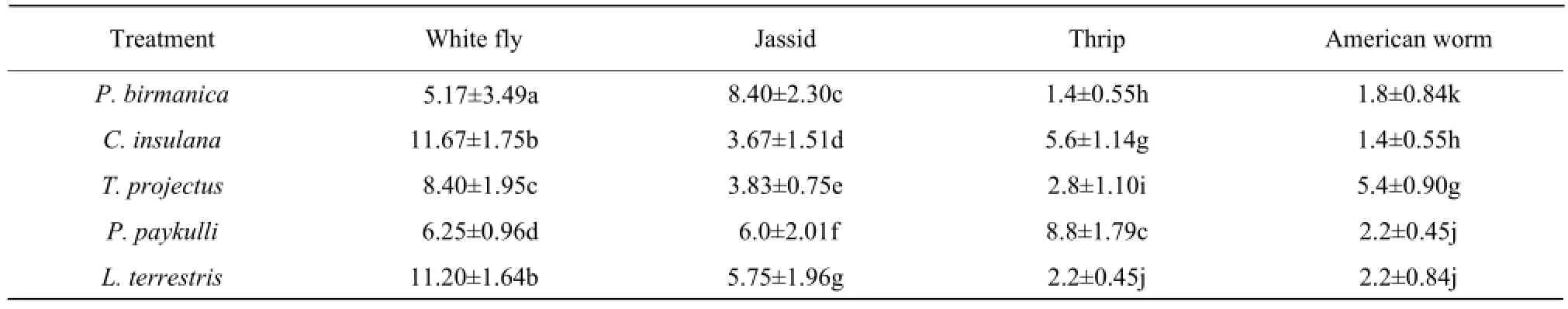
Table 2 Choice feeding of five male predatory spiders on different insect pests of cotton in choice chambers under control laboratory conditions
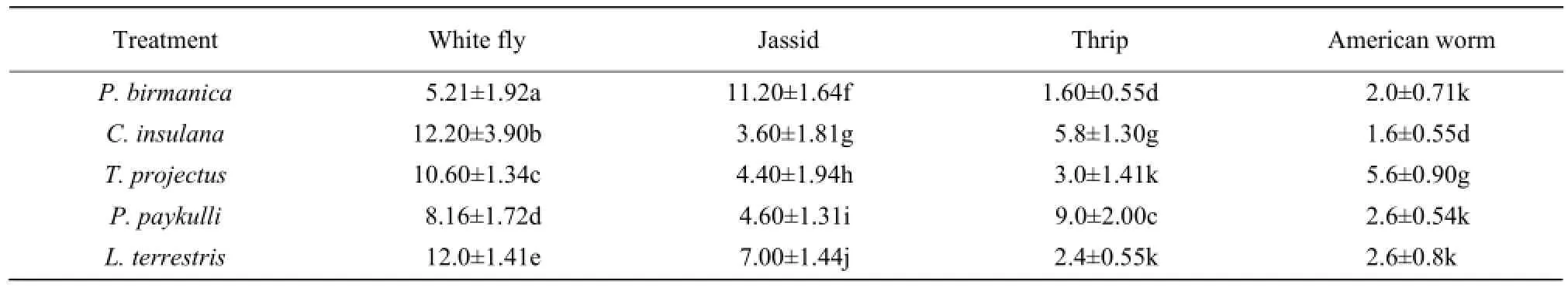
Table 3 Choice feeding of five female predatory spiders on different insect pests of cotton in choice chambers under control of laboratory conditions
It was found all the five spiders killed more the pests then feed and consumed only small portion of each pests. Mostly all the spiders were found to consuming the soft parts and suck the body fluid. So in the current study, the paralyzed or killed pests were considered as consumed pray. Fig. 2 depicted the percentage predation of each spider for both male and female.
No choice feeding
Results of no choice revealed that the predatory spiders consumed all the types of offered host pests (Table 4). Spiders showed different preferences to each host pest, but all the spider species showed higher efficacy of consuming the white fly (P. birmanica 10.40 ± 2.40, c. insulana 7.6 ± 1.51, T. projectus 8.1 ± 2.15, P. paykulli 8.2 ± 1.64 and L. terrestris 9.4 ± 1.94). Jassid was consumed more frequently by P. birmanica (10.20 ± 1.92) and the least by T. projectus (5.40 ± 1.51). Thrips was frequently predated by P. paykulli (8.6 ± 2.19) and least by P. birmanica (5.2 ± 1.30). All the spiders showed almost similar preference towards American bollworm with the highest preference of T. projectus (4.4 ± 0.55) and the least of P. paykulli (2.2±1.10).
Female spiders showed slightly higher predation in comparison of male perhaps of active nature of female spiders (Table 5). Fig. 3 represented higher percentage predation in case of no choice feeding because of nonavailability of other feeding resources and each spider was dependent on only one type of hosts. The spider being voracious predator killed most of the host pests in the no choice feeding.
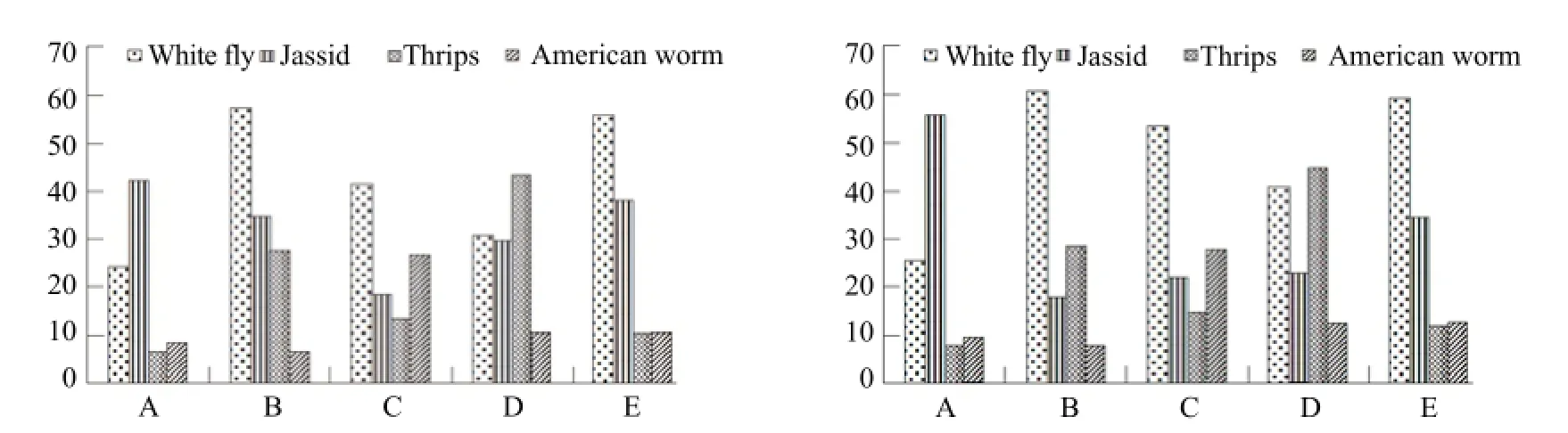
Fig. 2 Percentage consumption of male (left) and female (right) predatory spiders on four key cotton pests in choice feeding testA, P. birmanica; B, C. insuiana; C, T. projectus; D, P. paykulli; E, L. terrestris.

Table 4 No choice feeding of five female predatory spiders on different insect pests of cotton in choice chambers under control of laboratory conditions

Table 5 No choice feeding of five female predatory spiders on different insect pests of cotton in choice chambers under control of laboratory conditions

Fig. 3 Percentage consumption of male (left) and female (right) predatory spiders on four key cotton pests in no choice (single pest) feeding testA, P. birmanica; B, C. insuiana; C, T. projectus; D, P. paykulli; E, L. terrestris.
Discussion
Spiders were fed at different trophic levels and utilized herbivores detrivores and some other predators (Oelbemann et al., 2008). Being voracious and polyphagous predators, it utilized all the types of host pests offered to the spiders in the present study. Betterbody parts and better adaptations of prey capturing proved the spider a better predator in all the types of agro-ecosystems.
The pests killed in the no-choice feeding were higher as compared to the choice feeding due to the fact of non-availability of other feeding sources. Female consumed more prey in each set of experiment than male in similar conditions. This is due to the fact, female required more energies for egg laying, brood care and energy reservoir for reproduction and hibernation in the upcoming season (Walker and Rypstra, 2001). Moreover, the predation might also vary according to the life stages. Young spiders had better body parts and active in nature than sub-adults and aged spiders (Sebastian and Sudikumar, 2003). Among all the five species, L. terrestris showed slightly better predatory efficacy in both choice and no-choice feeding. This study was analogous to the study of Khuhro et al. (2012) who recorded predatory efficacy of seven predatory spiders in the cotton fields. They found P. birmanica actively consumed adult Jassid (11.80) than white fly and thrips. Our studies showed similar predation (11.20 ± 1.60) when 20 specimens were offered. The other similarity was the predation of T. projectus in case of Jassid, white fly and thrips. Khuhro et al. (2012) recorded T. projectus consumed 6.00 of Jassid, 5.60 of white fly and 3.20 of thrips when 20 samples of each pest were provided to each spider. Our study recorded 10.60 ± 1.34, 4.40 ± 1.94 and 3.0 ± 1.41 for white fly, Jassid and thrips, respectively. The difference of the predation might be due to fact, Khuhro et al. (2012) did not indicate whether they used the male or female spiders, but in our study, we used female spiders for the comparison with the study of Khuhro et al (2012). Overall the findings of this study revealed that all the predatory spiders consumed at least one pest host. This study supported the use of spiders in IPM and minimizing the uses and adverse effects of pesticides for future generations. The current investigated spiders might be strong candidates for controlling the cotton key pests like white fly, Jassid, thrips and bollworms.
Barrion A T, Litsinger J A. 1995. Riceland spiders of south and southeast Asia CAB International Wallingford UK, 701.
Butt A, Sherawat S M. 2012. Effect of different agricultural practices on spiders and their prey populations in small wheat fields. Acta Agric Scand section B- Soil Pl Sci,62: 374-382.
Channa S M, Kandhro K M M, Kumbhar R. 2006. Magnitude of gene action for different economic traits of upland cotton, Gossypiumhirsutum L. Pakistan J Agri, Agril Engg, Vet Sci,22(2): 16-20.
Gupta G P, Ajanta B, Seema R. 2004. Development of artificial diet for mass rearing of American bollworm, Helicoverpaarmigera. Indian Council of Agricultural Research,74(10): 548-551.
Hooks C R R, Pandey R R, Johnson M W. 2003. Impact of avian and arthropod predation on lepidopteran caterpillar densities and plant productivity in an ephemeral agroecosystem. Ecol Entomol,28: 522-532.
Jeyaparvathi S, Baskaran S, Bakavathiappan G A. 2013. Biological control potential of spiders on the selected cotton pests. International Journal of Pharmacy & Life Sciences,4(4): 2568-2572.
Khuhro R, Ghafoor A, Mahmood A, et al. 2012. Assessment of potential of predatory spiders in controlling the cotton Jassid (Amrascadevastans) under laboratory conditions. The Journal of Animal & Plant Sciences,22(3): 635-638.
Oelbermann K, Scheu S. 2009. Control of aphids on wheat by generalist predators: effects of predator density and the presence of alternative prey. Ent Exp Appl,132: 225-231.
Platnick. 2014. The world spider catalog, version 15. American Museum of Natural History. http://research.amnh.org/iz/spiders/catalog_15.0/ COUNTS.html.
Rajeswaran J, Duraimurugan P, Shanmugam P S. 2005. Role of spiders in agriculture and horticulture ecosystem. Food Agriculture & Environment (JFAE),3(3/4): 147-152.
Sebastian P A, Sudhikumar A V. 2003. Feeding potential of spiders (Order: Araneae) on Aphis craccivora Koch occurring on cotton Entomon,28(2): 153-156.
Sharma S. 2014. A study on spiders as predators in the agro-ecosystems. Munis Entomology & Zoology,9(1): 80-83.
Tikader B K, Biswas B. 1981. Spider fauna of Calcutta and vicinity. Part I, Rec Zool Surv India Pub,30: 1-148
Walker E S, Rypstra L A. 2001. Sexual dimorphism in functional response and troph morphology in Rabidosarabida (Araneae: Lycosidae). Am Midl Nat,146: 161-170.
Q96
A
1006-8104(2015)-03-0048-06
Received 16 June 2015
Mohsin S B (1984-), male, Ph. D, engaged in the research of preventive veterinary medicine. E-mail: mohsinbukhari70@gmail.com
 Journal of Northeast Agricultural University(English Edition)2015年3期
Journal of Northeast Agricultural University(English Edition)2015年3期
- Journal of Northeast Agricultural University(English Edition)的其它文章
- Design of Non-contact On-load Automatic Regulating Voltage Transformer
- Chinese Comprehensive Rural Reform: Institutional Vicissitude, Theoretic Framework and Content Structure
- Acceptability of Bush Meat as a Source of Animal Protein in Delta State, Nigeria: Implication for Extension Services
- Preparation of Mouse Embryonic Stem Cells and Cardiomyocyte Differentiation Induced with Retinoic Acid and Ascorbic Acid
- Contents of Trace Metal Elements in Cow Milk Impacted by Different Feedstuffs
- Effect of Bacillus subtilis and Pseudomonas fluorescens on Growth of Greenhouse Tomato and Rhizosphere Microbial Community